Physical Address
304 North Cardinal St.
Dorchester Center, MA 02124

All lower motor neuron (LMN) groups except the facial nerve nucleus that supplies the muscles of facial expression consist of both alpha LMNs that supply the skeletal muscle fibers (extrafusal fibers) and gamma LMNs that supply the small contractile elements in the muscle spindles (intrafusal fibers). The muscles of facial expression do not have muscle spindles and are not supplied by gamma LMNs. The alpha LMNs regulate contraction of the skeletal muscles to produce movement. The gamma LMNs regulate the sensitivity of the muscle spindles for group Ia and group II afferent modulation of alpha LMN excitability.
An alpha LMN supplies a motor axon to a variable number of skeletal muscle fibers (extrafusal fibers), ranging from just a few (e.g., extraocular muscles) to several thousand (large muscles such as the quadriceps). The LMN and its innervated skeletal muscle fibers are called a motor unit . Supporting cells (such as Schwann cells) and myocytes produce trophic factors to maintain the nerve-muscle association; when nerve injury occurs, growth factors help to attract motor axonal regrowth to reestablish the prior nerve-muscle association. When motor axons degenerate, the neuromuscular junctions (NMJs) disappear, and the nicotinic cholinergic receptors spread across the membrane of the denervated skeletal muscle fibers. This results in denervation hypersensitivity to nicotinic cholinergic stimulation, noted as random individual muscle fiber twitches (fibrillation), best observed by electromyography. If motor nerves are attracted back to the muscle fibers and NMJs are restored, the nicotinic cholinergic receptors are again restricted to the secondary folds of the NMJ. If the motor axon that was lost cannot regrow, neighboring motor axons of other motor units that supply adjacent skeletal muscle fibers may send sprouts to the denervated muscle fibers and incorporate them into the motor unit; the consequence is a larger motor unit and a greater demand on the LMN cell body that now supplies a greater than normal number of skeletal muscle fibers. This mechanism may account for recovery of physiological function in some LMN diseases such as polio. If the alpha-LMN cell body itself is damaged or is in the process of dying (e.g., in amyotrophic lateral sclerosis), the axon may produce aberrant action potentials (agonal bursts of electrical activity) that result in muscle fiber contraction throughout the motor unit, called a fasciculation, which is visually observable. A denervated muscle fiber must be reinnervated within 1 year or so if it is to restore relatively normal function; a longer period leads to permanent changes that preclude proper reinnervation. Many experimental approaches are seeking to restore innervation or attract a more robust nerve supply to denervated muscle fibers by applying or inducing gene expression of growth factors and trophic factors. Denervated skeletal muscle fibers are flaccidly paralyzed, lack muscle tone, cannot be induced to contract with muscle stretch reflexes, and undergo atrophy; these are classic characteristics of LMN syndrome .
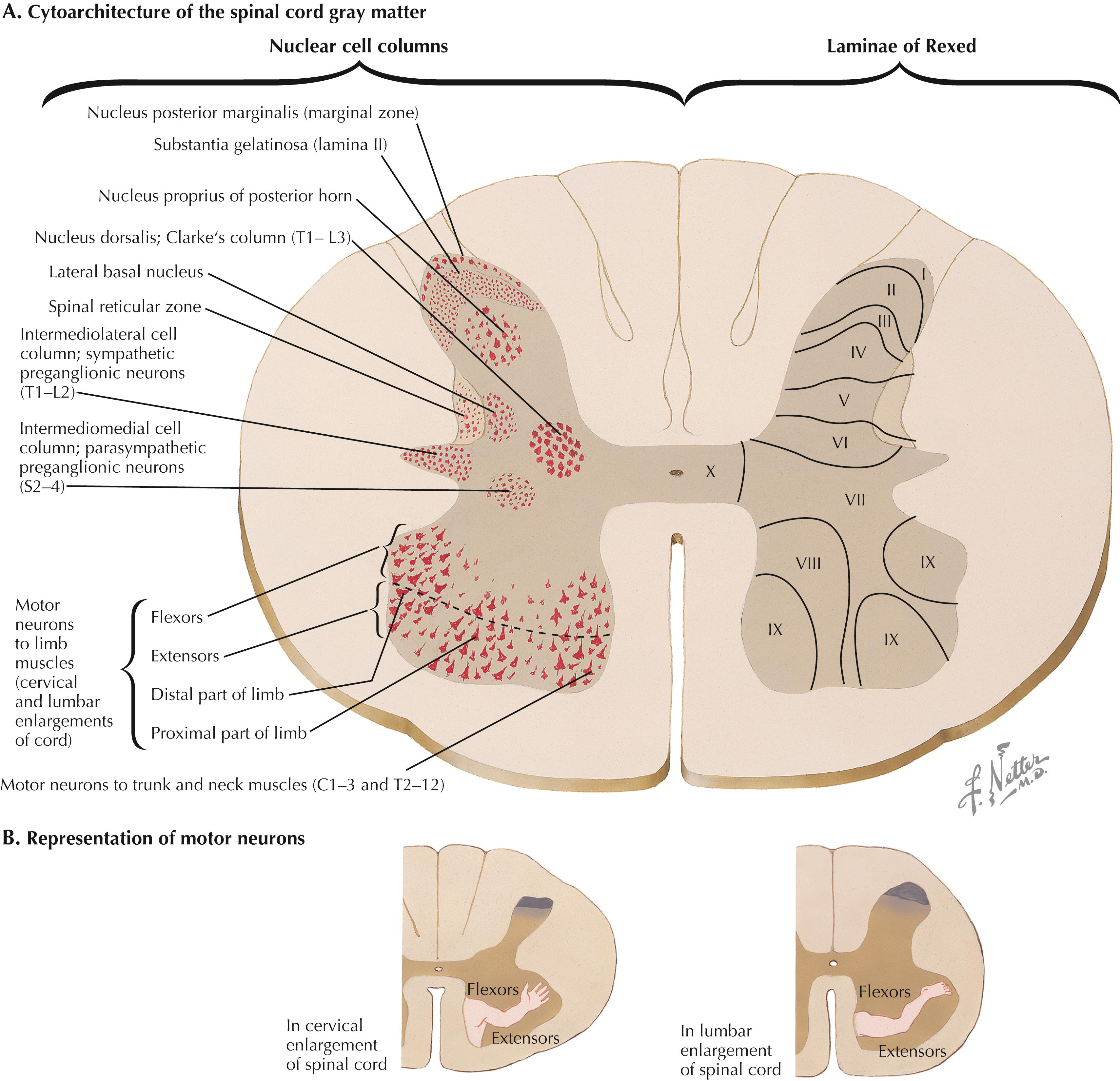
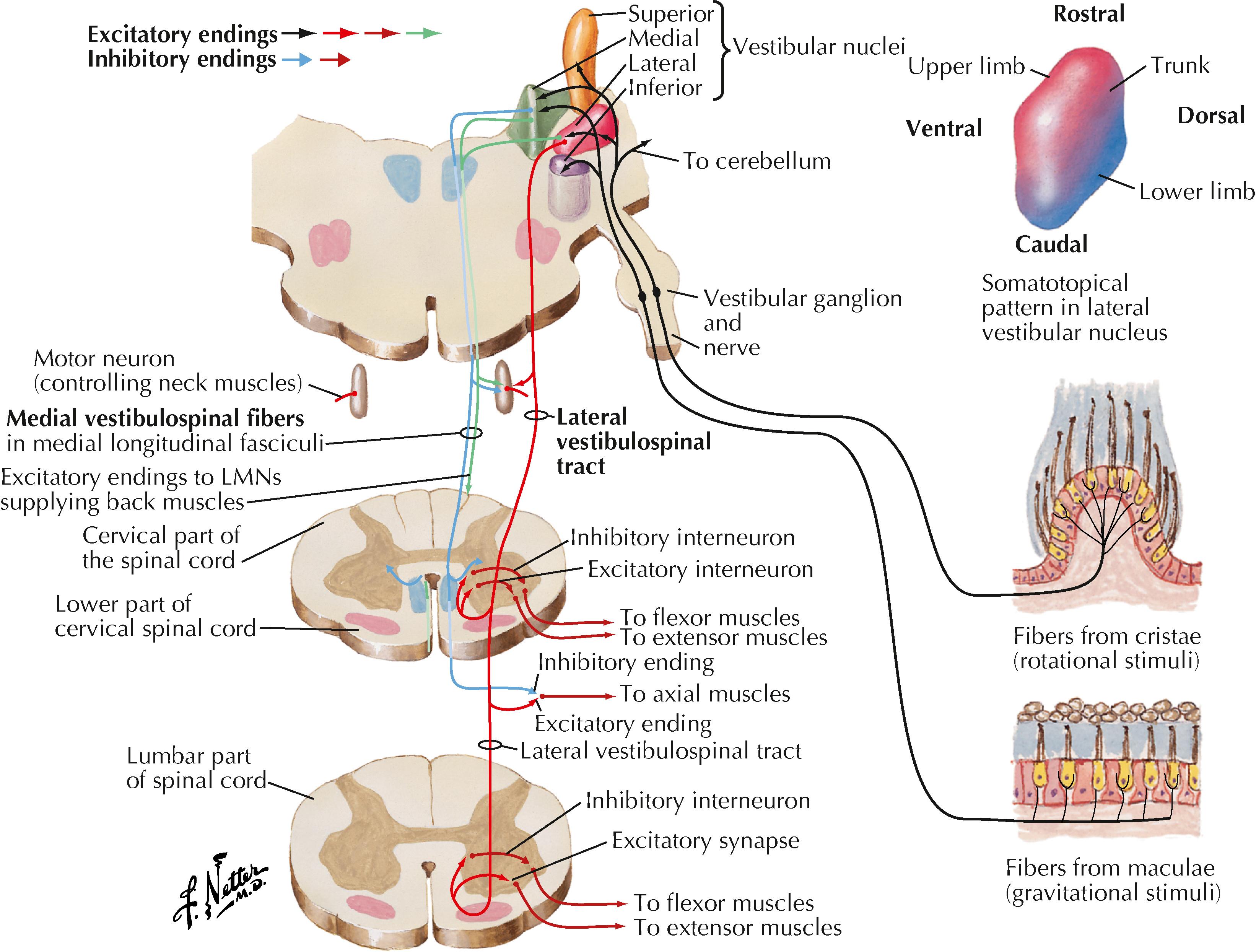
LMNs are found as clusters of neurons in the anterior (ventral) horn of the spinal cord, represented as lamina IX of Rexed. Distinct clusters of LMNs supply distinct skeletal muscles with motor innervation. These LMN groups are organized topographically; LMNs distributing to trunk and neck muscles are found medially, and LMNs distributing to muscles of distal extremities are found laterally. Within spinal cord segments, LMNs distributing to flexor muscle groups are found dorsally, and LMNs distributing to extensor muscle groups are found ventrally.
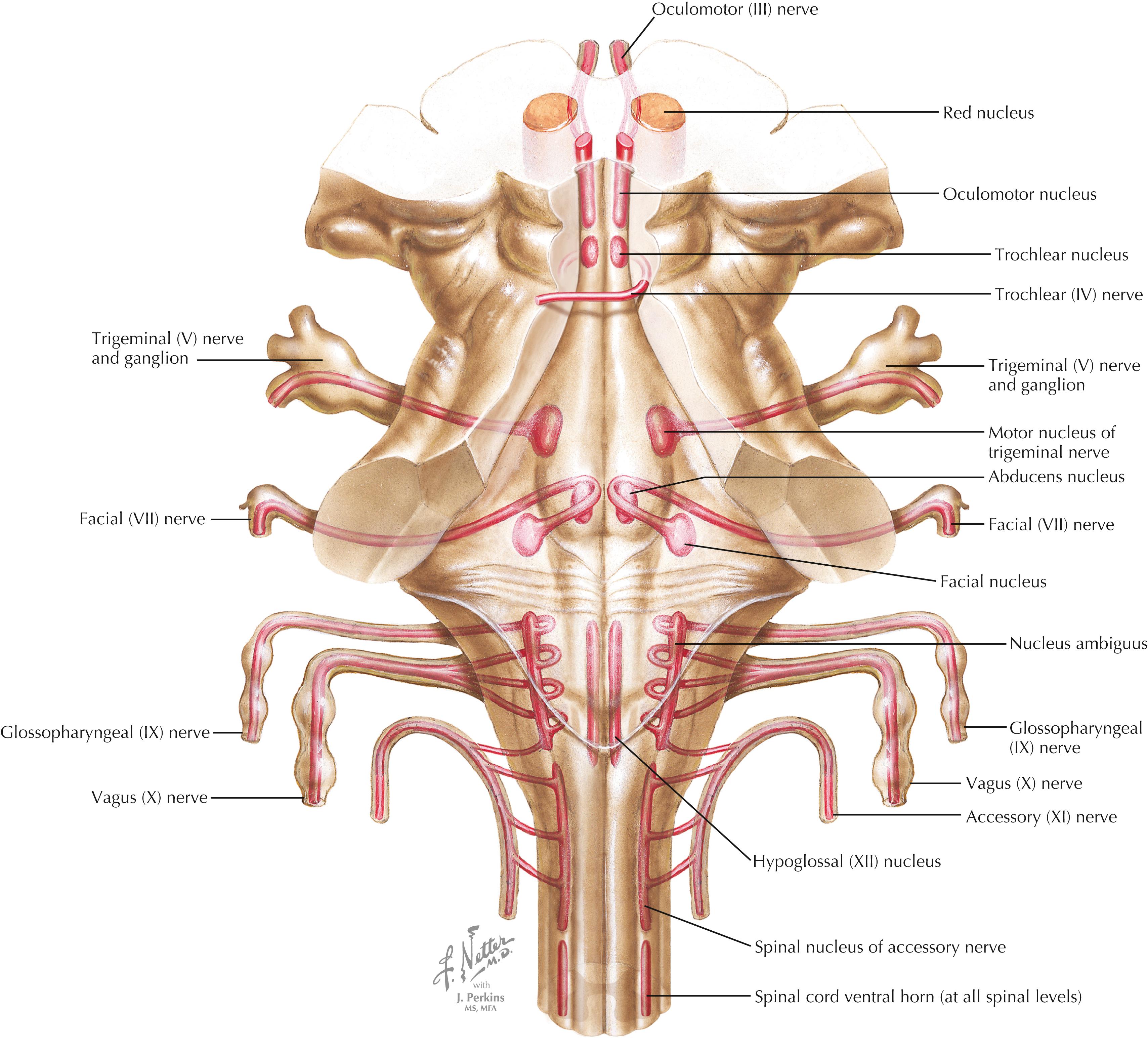
LMNs are found in medial and lateral columns in a longitudinal view of the brainstem. The medial column (LMNs of the oculomotor nucleus, trochlear nucleus, abducens nucleus, and hypoglossal nucleus) derives from the general somatic efferent system, and the lateral column (LMNs of motor nucleus V, facial nucleus, nucleus ambiguus, and spinal accessory nucleus) derives from the special visceral efferent system. LMNs in the spinal cord are found in a longitudinal column coursing through the anterior horn at all levels.
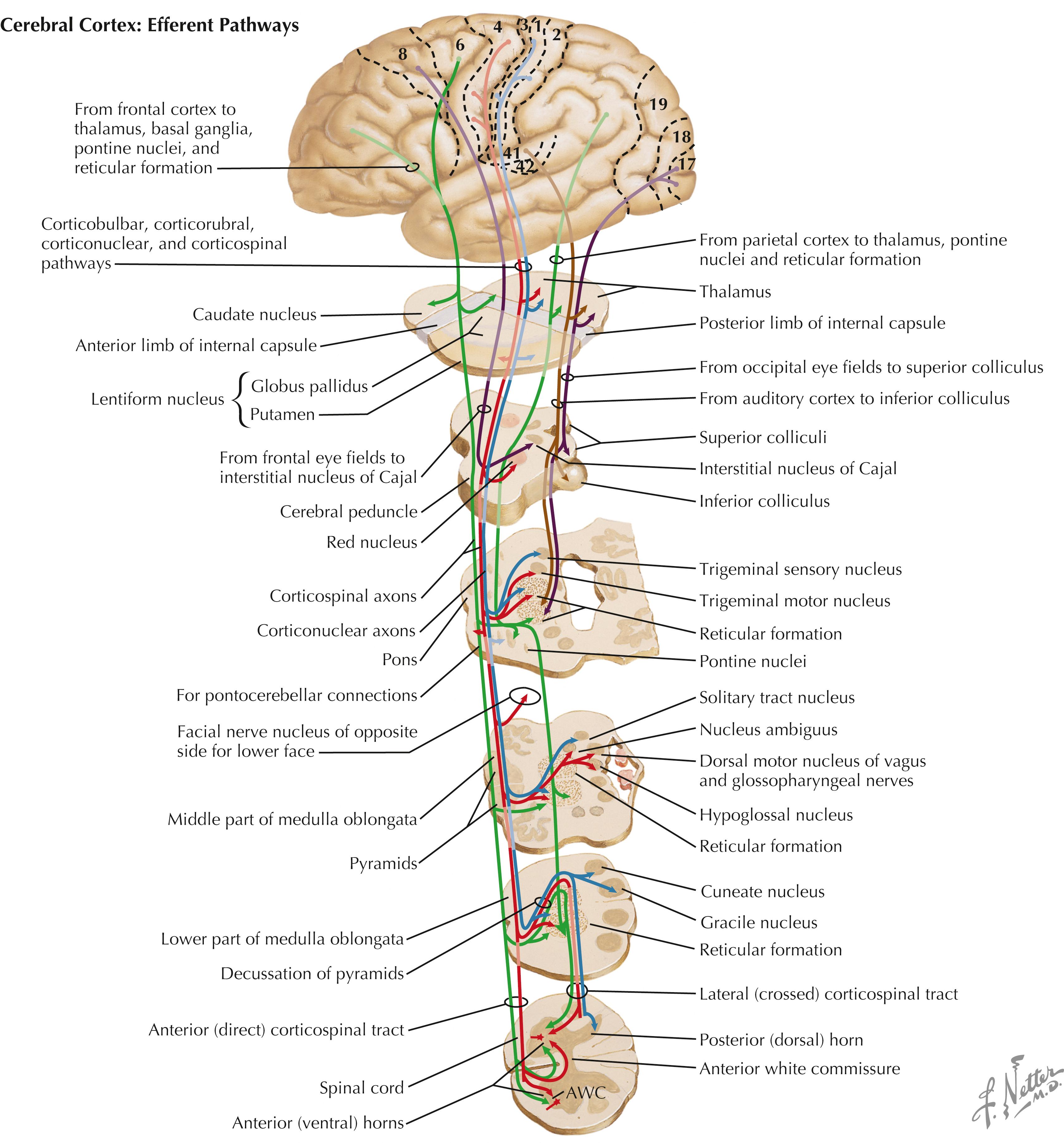
Cortical neurons in the motor cortex (area 4) and the supplemental and premotor cortices (area 6) send axons to the basal ganglia (caudate nucleus and putamen), the thalamus (ventral anterior [VA] and ventral lateral [VL] nuclei), the red nucleus, the pontine nuclei, the cranial nerve (CN) motor nuclei on both sides, and the spinal cord ventral horn, mainly on the contralateral side. These axons form the corticospinal tract, corticobulbar tract, corticostriatal projections, corticopontine projections, corticothalamic projections, and cortical connections to the upper motor neurons (UMNs) of the brainstem (reticular formation [RF] motor areas, red nucleus, superior colliculus). Neurons of the sensory cortex (areas 3, 1, 2) send axons mainly to secondary sensory nuclei (corticonuclear fibers) to regulate incoming lemniscal sensory projections destined for conscious interpretation. Neurons in the frontal eye fields (area 8) project to the superior colliculus, the horizontal and vertical gaze centers of the brainstem, and the interstitial nucleus of Cajal to coordinate voluntary eye movements and associated head movements. Other regions of sensory cortex project axons to thalamic and brainstem structures that regulate incoming lemniscal sensory information. Some cortical efferent fibers project to limbic forebrain regions, such as the amygdaloid nuclei, hippocampal formation, and septal nuclei.
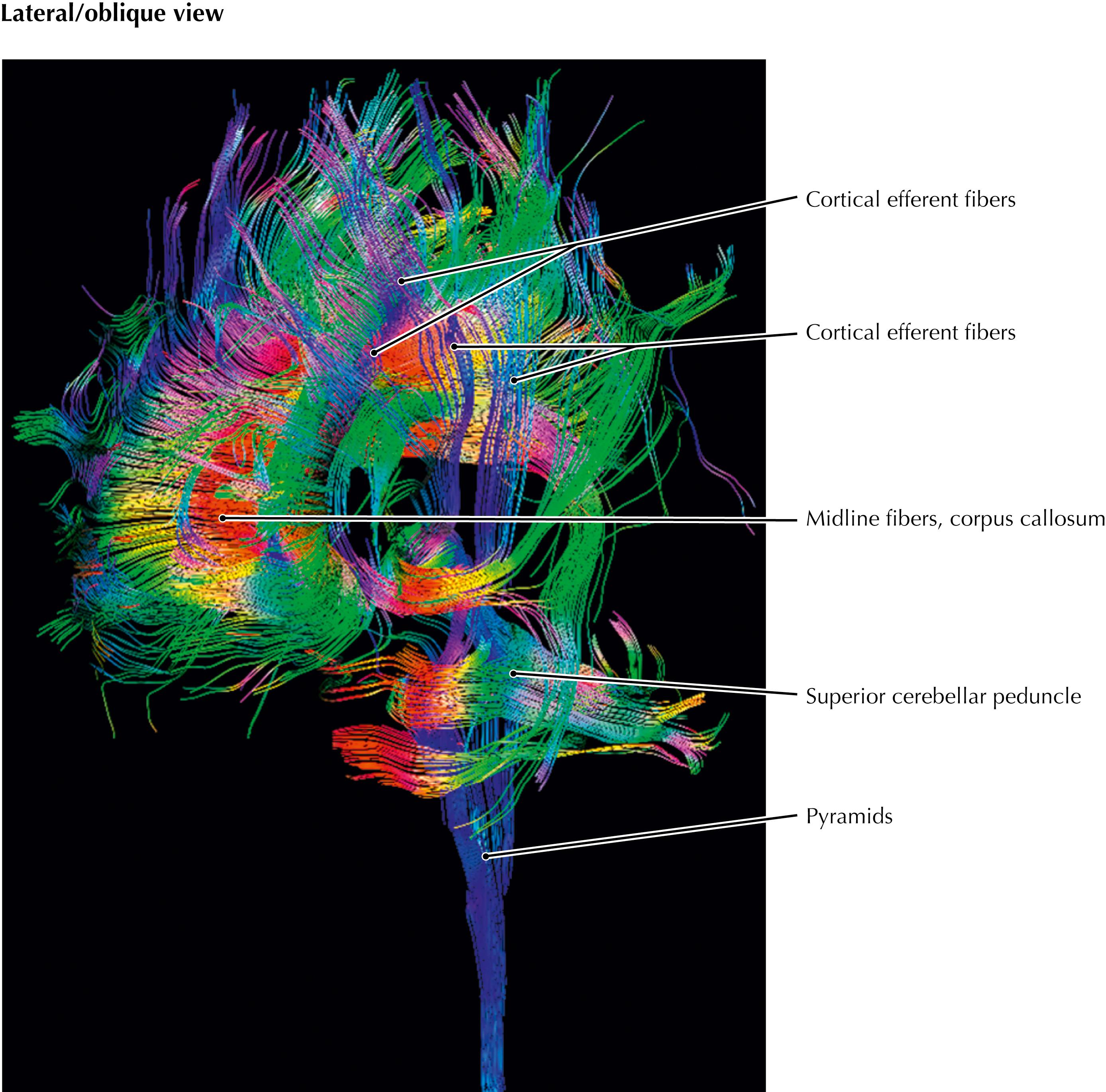
This diffusion tensor image shows the cortical efferent pathways in a lateral oblique section. These pathways, shown in blue, channel from widespread areas of the cerebral cortex to structures in the forebrain, the thalamus, the brainstem, the cerebellum (indirectly, through the pontine nuclei), and the spinal cord. Additional cortical association pathways are depicted in green (running in anterior-posterior direction) and commissural pathways are shown in red (running in left-right direction).
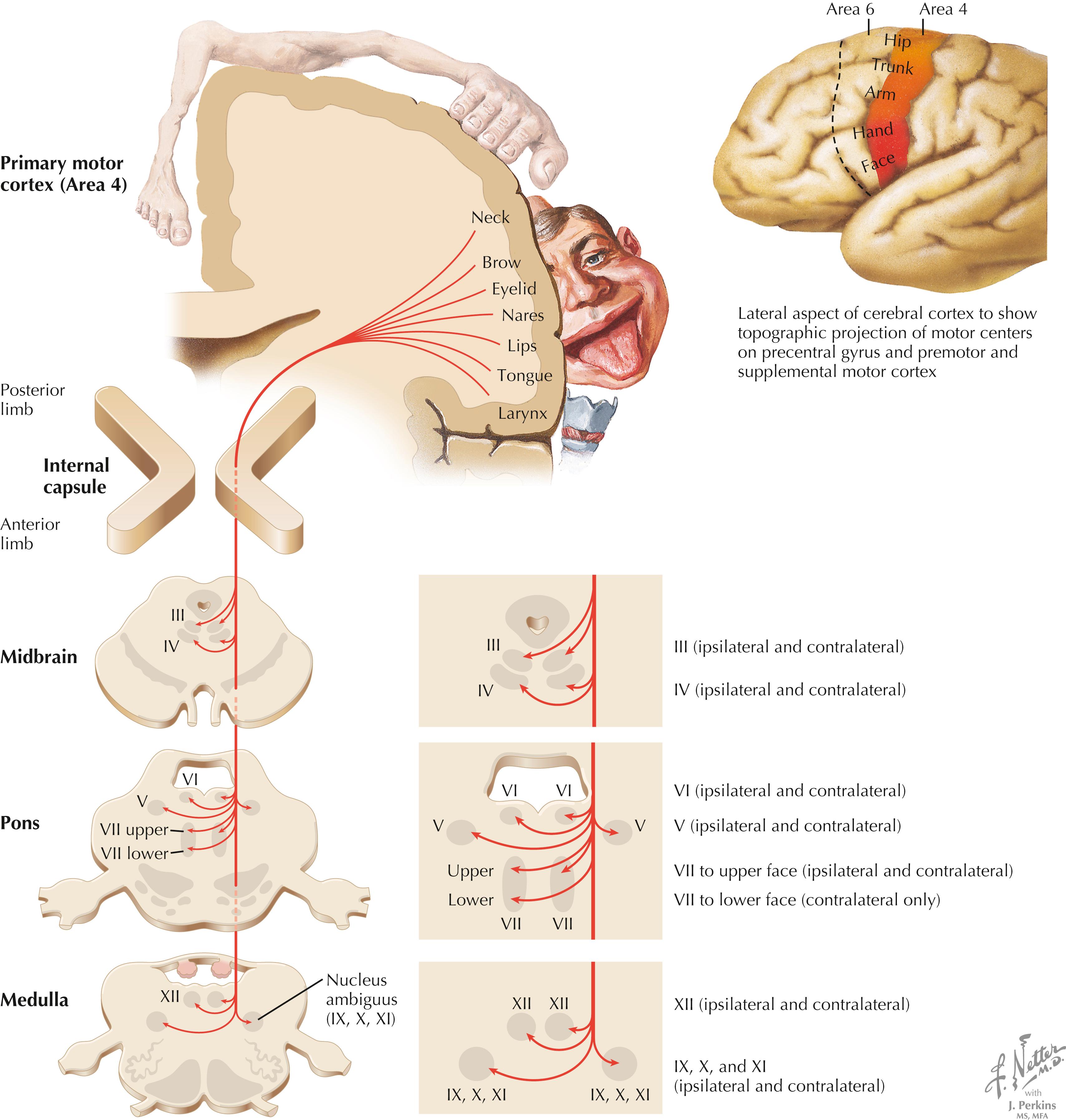
The corticobulbar tract (CBT) arises mainly from the lateral portion of the primary motor cortex (area 4). CBT axons project through the genu of the internal capsule into the cerebral peduncle, the basis pontis, and the medullary pyramids on the ipsilateral side. The axons distribute to CN motor nuclei on the ipsilateral and contralateral sides except for the portion of the facial nerve nucleus (CN VII) that supplies the muscles of facial expression for the lower face, which receives exclusively contralateral projections. The CBT projections to the hypoglossal nucleus are mainly contralateral; CBT projections to the spinal accessory nucleus are mainly ipsilateral. CBT lesions result mainly in contralateral drooping of the lower face that is paretic to attempted movements from voluntary commands (central facial palsy), in contrast to Bell’s palsy (CN VII palsy), in which the entire ipsilateral face is paralyzed.
See next page.
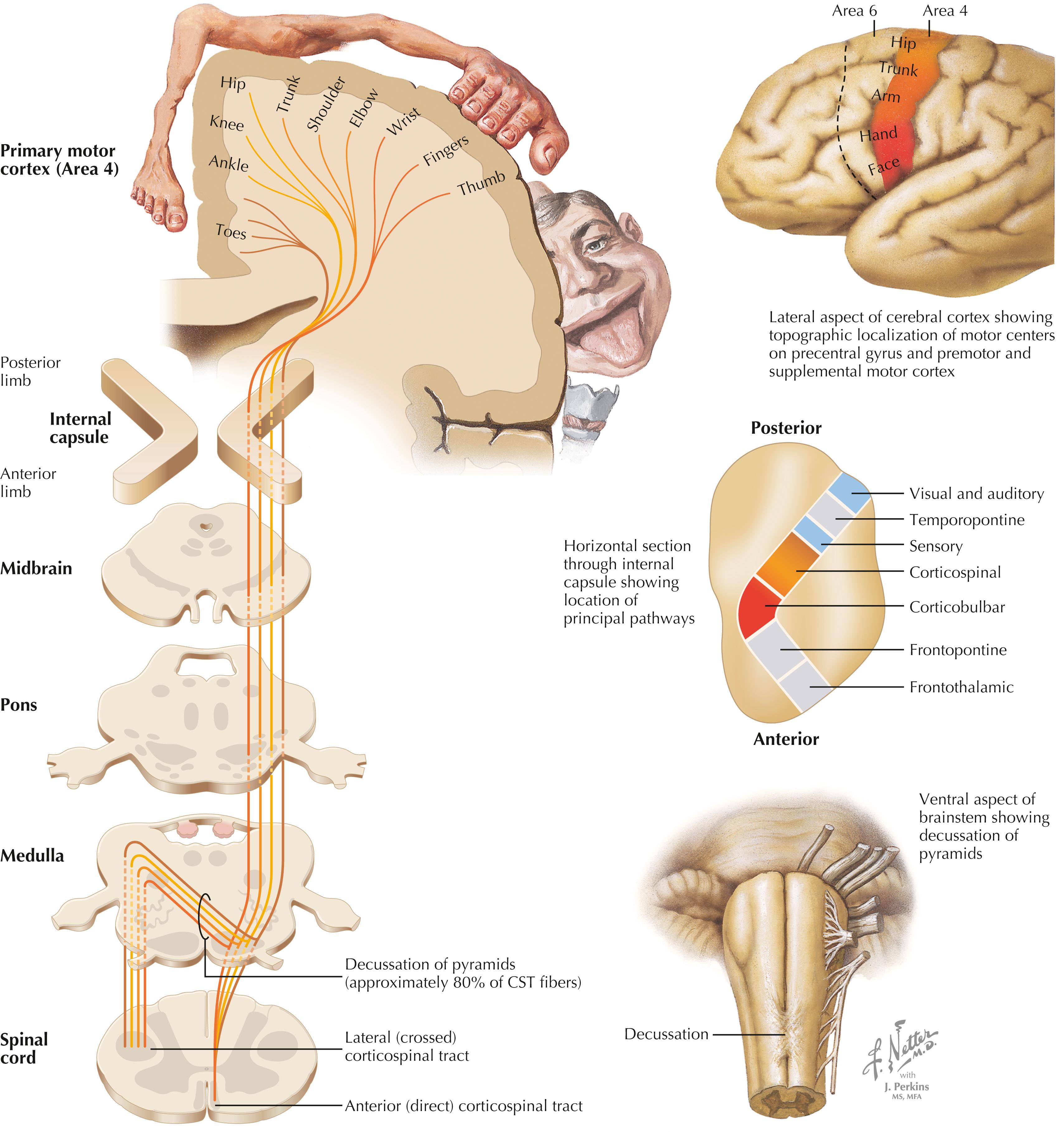
The motor portion of the corticospinal tract (CST) originates from neurons of many sizes, mainly from the primary motor cortex (area 4) and the supplemental and premotor cortices (area 6). The primary sensory cortex (areas 3, 1, 2) contributes axons into the CST, but these axons terminate mainly in secondary sensory nuclei to regulate the processing of incoming lemniscal sensory information. The CST travels through the posterior limb of the internal capsule, the middle region of the cerebral peduncle, numerous fascicles of axons in the basis pontis, and the medullary pyramid on the ipsilateral side. Most of the CST axons (approximately 80% but variable from individual to individual) cross the midline in the decussation of the pyramids at the medullary–spinal cord junction. These crossed fibers descend in the lateral CST in the lateral funiculus of the spinal cord and synapse on alpha and gamma LMNs, both directly and indirectly through interneurons. CST axons that do not decussate continue as the anterior CST in the anterior funiculus of the spinal cord and then decussate at the appropriate level through the anterior white commissure to terminate directly and indirectly on alpha and gamma LMNs contralateral to the cortical cells of origin. Only a very small portion of the motor connections of the corticospinal tract terminate on LMNs on the ipsilateral side of the spinal cord.
The motor portion of the CST arises mainly from neurons in the primary motor cortex (area 4) and the supplemental and premotor cortices (area 6). The primary sensory cortex and superior parietal lobule contribute corticospinal axons (corticonuclear fibers) to secondary sensory nuclei in the lower brainstem and spinal cord. Approximately 80% of the CST axons cross in the decussation of the pyramids and terminate directly and indirectly with alpha and gamma LMNs that control movements of the distal extremities, especially the hands and fingers. At least 10% of the CST terminates monosynaptically on alpha LMNs, especially those associated with hand and finger musculature. A lesion in the internal capsule damages the CST, corticorubral fibers, and corticoreticular fibers, resulting in contralateral hemiplegia. Initially, the hemiplegia is flaccid, with loss of tone and reflexes. Within days to a week or so, the hemiplegia becomes spastic, with hyperreflexia and hypertonus. The affected musculature shows initial resistance to attempted passive movement, followed by a dissipation or “melting” of tone (the clasp-knife reflex), perhaps because of high threshold Ib Golgi tendon organ inhibitory influences on the homonymous LMNs. The initial suspected mechanism of classical UMN syndrome was disinhibition of dynamic gamma LMNs, which drives initial resistance to passive stretch, mediated via subsequent Ia afferent influences over alpha LMNs; this mechanism was reinforced by observations that dorsal root sectioning diminished spasticity in UMN syndromes. Further studies have revealed additional potential mechanisms, including diminished reciprocal inhibition, recurrent Renshaw inhibition, and presynaptic inhibition on Ia afferents, all suggestive of major changes in interneurons of the spinal cord following a classic UMN lesion. In UMN syndrome, the plantar reflexes are extensor (reverting to a developmentally early stage in the absence of the CST), and abdominal reflexes are absent on the affected side. Clonus (repetitive alternating flexor and extensor muscle stretch reflexes) also may occur and is possibly attributable to interneuronal changes such as diminished Renshaw inhibition.
The CBT arises mainly from the lateral portion of the primary motor cortex; it descends through the genu of the internal capsule and the cerebral peduncle (medial to the corticospinal tract fibers) ipsilaterally, and it distributes bilaterally to the motor CN nuclei (CNN) of the brainstem, except to the facial nucleus for the lower face, which receives almost exclusively contralateral projections. The cortico-bulbar axons terminate mainly on interneurons that regulate LMN output. Originally, corticobulbar was a term reserved for cortical projections to LMNs of the medulla (bulb), but it now has been expanded to include CNN for V, VII, nucleus ambiguus, XII, and the spinal accessory (XI) nucleus. A lesion in the genu of the internal capsule (embolic or thrombotic stroke or hemorrhage of the middle cerebral artery or its branches) or the cerebral peduncle (Weber’s syndrome, compression of the peduncle against the free edge of the tentorium cerebelli with transtentorial herniation) results mainly in a drooping lower face (central facial palsy) on the contralateral side. The intact hemisphere can control voluntary movement of the LMNs in the CNN for all other brainstem motor nuclei on both sides. In some individuals, a predominance of contralateral fibers to LMNs for the soft palate or the tongue is noted, resulting in a temporary contralateral palsy, or a predominance of ipsilateral fibers to LMNs of XI may be noted, resulting in an ipsilateral palsy of the sternocleidomastoid and upper trapezius muscles. This central paresis occurs without atrophy. Bilateral corticobulbar lesions result in profound paralysis of voluntary movement in all muscles supplied by CNN, with preservation of muscle bulk, reflex responses, and some emotional responses using those LMNs. The LMNs in CNN III, IV, and VI receive cortical input from the frontal eye fields (area 8) and parietal eye fields of both sides.
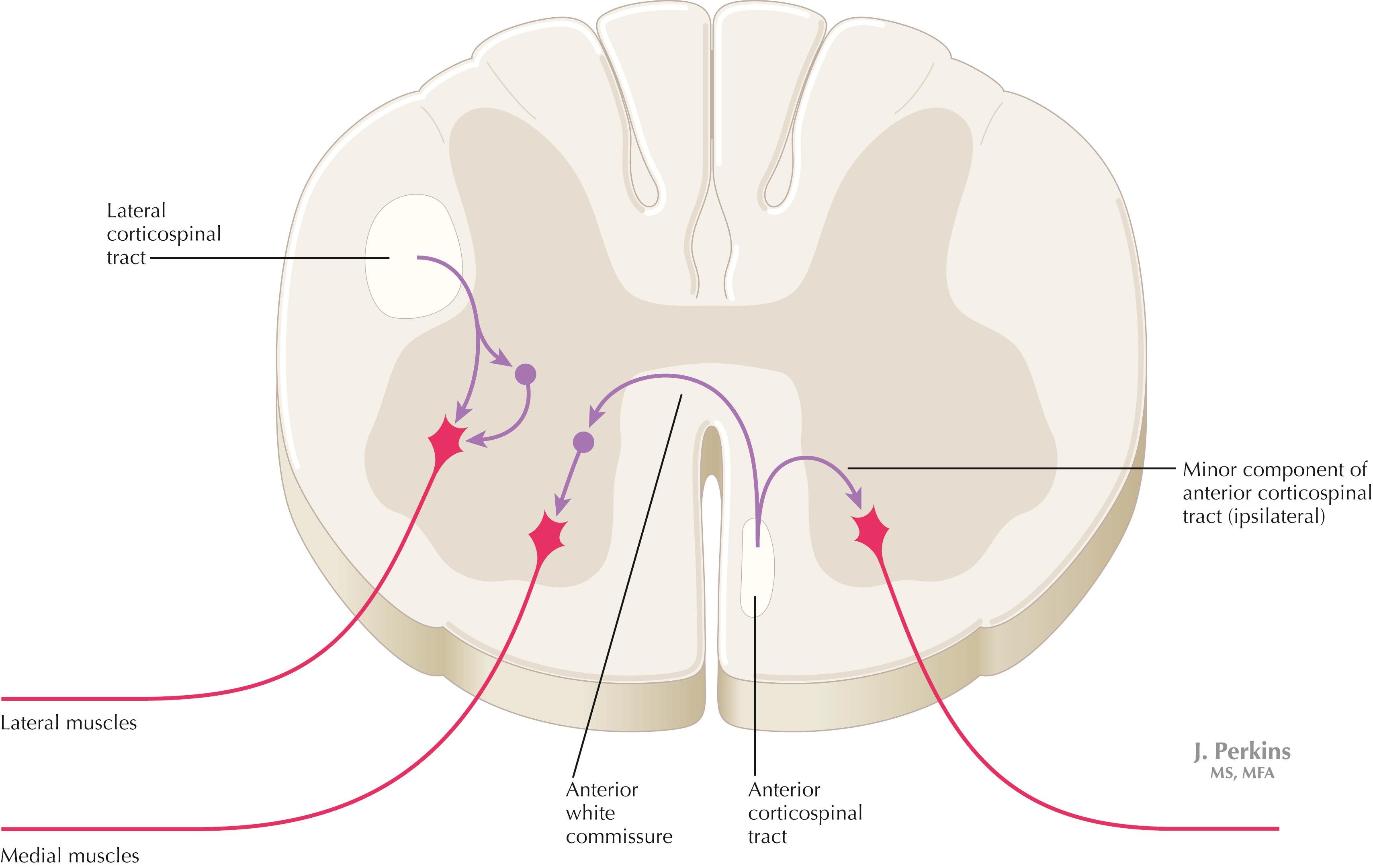
Crossed axons in the lateral CST, intermixed with axons of the rubrospinal tract, travel in the lateral funiculus. These CST axons terminate directly and indirectly mainly on LMNs associated with distal musculature, especially for skilled hand and finger movements. The uncrossed anterior CST axons decussate predominantly in the anterior white commissure and terminate directly and indirectly mainly on LMNs that supply medial musculature. A small number of anterior CST axons terminate ipsilateral to the cortical cells of origin. An isolated lesion in the CST in the medullary pyramids results in weakness of contralateral fine, dexterous hand and finger movements. All other lesions involving the CST at other levels (internal capsule, cerebral peduncle, pons), where these descending fibers are intermixed with other descending motor systems, produce contralateral spastic hemiplegia with hypertonus, hyperreflexia, and plantar extensor responses as long-term consequences. Lesions in the lateral CST produce similar symptoms ipsilateral to the damaged lateral funiculus below the level of the lesion.
Cerebral palsy is a general term referring to a group of motor impairment syndromes from congenital or developmental lesions. Approximately two-thirds of these lesions occur prenatally, and 10% occur perinatally. Some cases appear as delays in developmental milestones.
The motor deficits may include monoplegia, diplegia, hemiplegia, or quadriplegia. The most common appearance (75%) is lower extremity diplegia with scissoring gait (adductor spasms), sometimes accompanied by hip, knee, or elbow flexion. Hemiplegic cerebral palsy mainly affects the upper extremity, accompanied by a persistent grasp reflex. Motor characteristics often are spastic but may be hypotonic or atonic, or dyskinetic. Quadriplegic cerebral palsy is often accompanied by cortical defects, mental impairment, seizures, visual defects, and oculomotor defects.
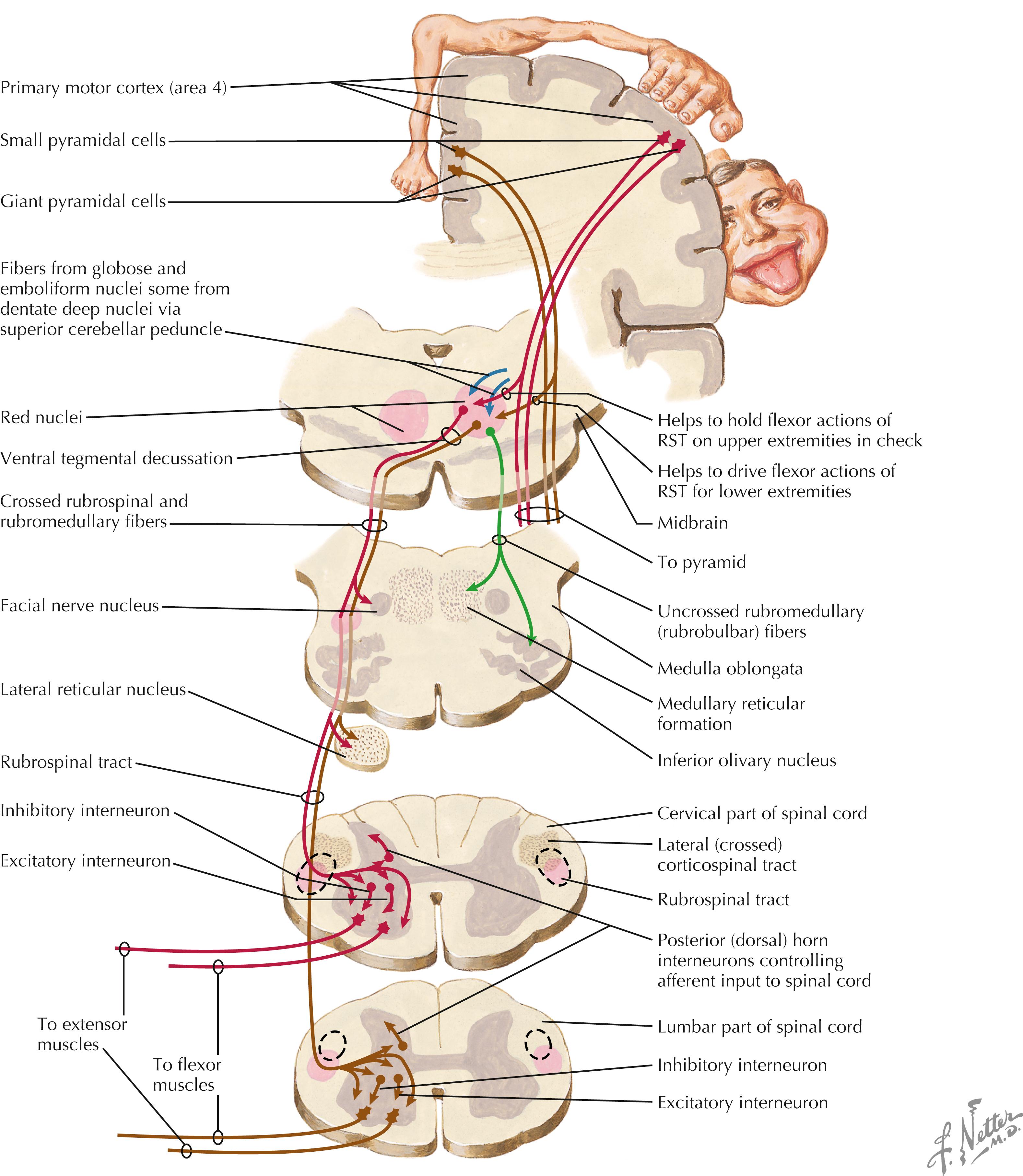
The cortico-rubro-spinal system is an indirect corticospinal system that regulates spinal cord LMNs. The red nucleus in the midbrain receives topographically organized ipsilateral connections from the primary motor cortex (area 4). Axons of the rubrospinal tract (RST) decussate in the ventral tegmental decussation and descend in the lateral brainstem and the lateral funiculus of the spinal cord, where they are intermixed extensively with axons of the lateral CST. The RST terminates directly and indirectly on alpha and gamma LMNs in the spinal cord, particularly those associated with flexor movements of the extremities. The RST helps to drive flexor movements of the upper extremity and helps to hold in check flexor movements of the lower extremity. RST lesions usually occur in conjunction with the CST in the spinal cord; corticorubral lesions also occur in conjunction with the CST in the internal capsule and cerebral peduncle. These lesions result in contralateral spastic hemiplegia as long-term consequences. Brainstem lesions caudal to the red nucleus result in decerebration (extensor spasticity), reflecting the removal of the flexor drive of the rubrospinal tract to LMNs supplying the upper limbs. See page 433 for a Clinical Point.
Become a Clinical Tree membership for Full access and enjoy Unlimited articles
If you are a member. Log in here