Physical Address
304 North Cardinal St.
Dorchester Center, MA 02124

The neural plate, neural tube, and neural crest form at the 18-day stage of embryonic development. The underlying notochord induces the neural plate, and a midline neural groove forms. The elevated lateral margins become the neural folds, tissue destined to become the neural crest with future contributions to many components of the peripheral nervous system (PNS). At this very early stage of embryonic development, these neural precursors are vulnerable to toxic insult and other forms of damage.

In the 21– or 22-day-old-embryo, the neural plate, with its midline neural groove, thickens and begins to fold and elevate along either side, allowing the two lateral edges to fuse at the dorsal midline to form the completed neural tube. The central canal, the site of the future development of the ventricular system, is in the center of the neural tube. This process of neurulation continues both caudally and rostrally. Disruption can occur because of failure of full neural tube formation caudally (spina bifida) or rostrally (anencephaly).
As the neural plate forms into a neural tube, the process of neurulation results in fused neural folds, starting centrally and moving both caudally and rostrally. Failure of the neural tube to close results in dysraphic defects, with altered development of associated muscles, bone, skin, and meninges. If the anterior neuropore fails to form, anencephaly results, with failure of the brain to develop, accompanied by facial defects. This condition is lethal. Failure of the posterior (caudal) neuropore to close results in spina bifida , with failure of the vertebral arches to fuse. A saccular protrusion from the lumbar region may contain meninges (meningocele) or meninges and spinal cord (meningomyelocele). Meningomyelocele is often accompanied by paraparesis, bowel and bladder dysfunction, sensory disruption at the level of the lesion, motor dysfunction in the lower extremities, and accompanying hydrocephalus or Arnold-Chiari malformation, requiring a ventriculo-peritoneal or ventriculo-jugular shunt.
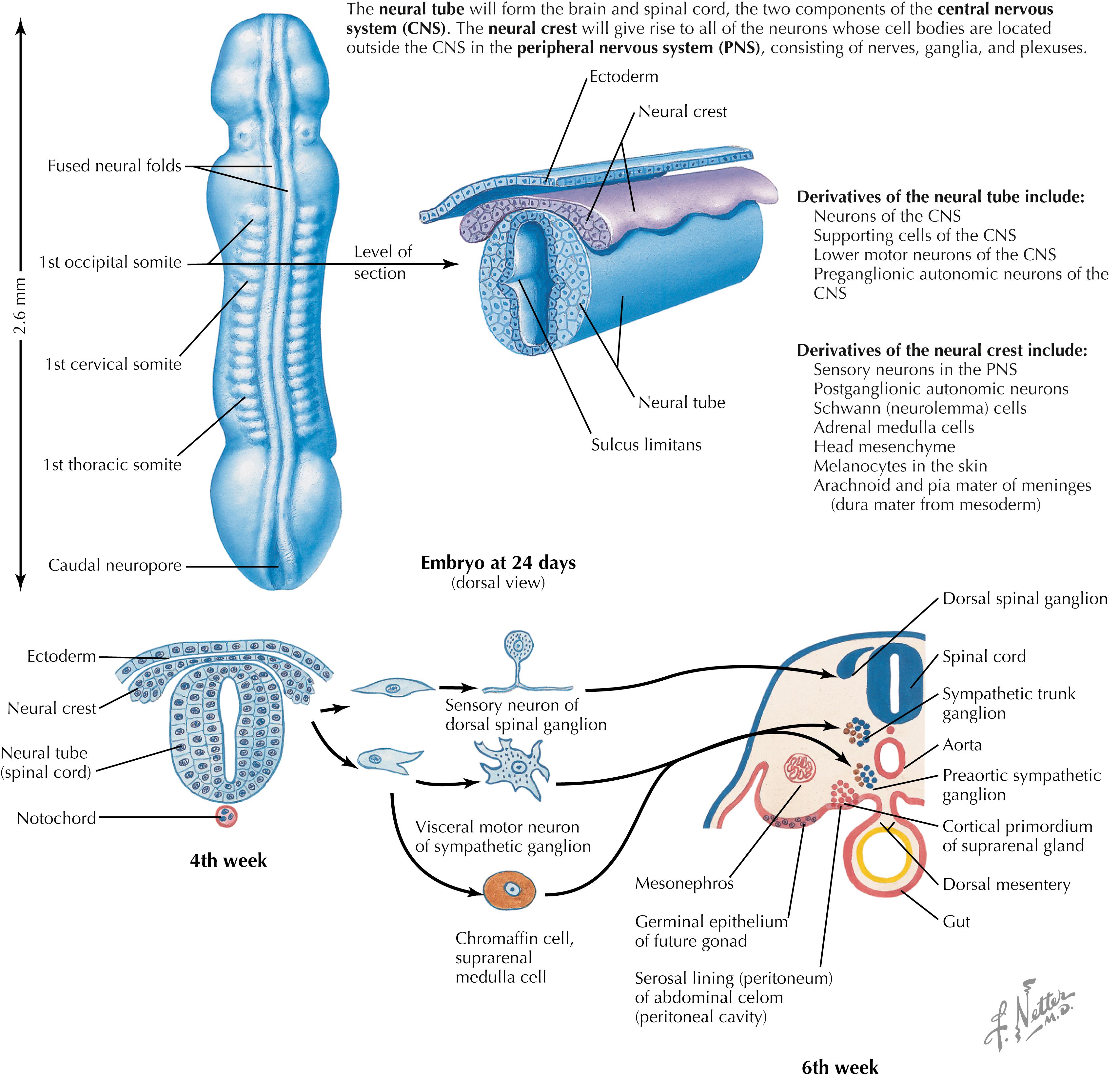
The dorsal and ventral halves of the neural tube are separated by the sulcus limitans, an external protrusion from the central canal that demarcates the alar plate above from the basal plate below. This important landmark persists at some sites in the adult ventricular system. The alar plate is the source of generation of many neurons with sensory function. The basal plate is the source of generation of many neurons with motor or autonomic function in the spinal cord and the brainstem. The neural crest cells at the edge of the neural folds unite and become a dorsal crest, with the neural crest above the neural tube. The neural tube and neural crest separate from the originating ectoderm.
The neural crest gives rise to a wide variety of neural elements of the PNS, including primary sensory neurons, postganglionic autonomic neurons, Schwann cells, adrenal medullary chromaffin cells, pia and arachnoid cells, melanocytes, and some mesenchyme of the head. A failure of the neural crest to develop and migrate properly is seen in Hirschsprung’s disease (congenital megacolon), in which sensory signals from the colon are absent, and in familial dysautonomia, in which autonomic symptoms (cardiovascular dysfunction, gastrointestinal dysfunction) and sensory deficits (especially pain and temperature sensation) are present.
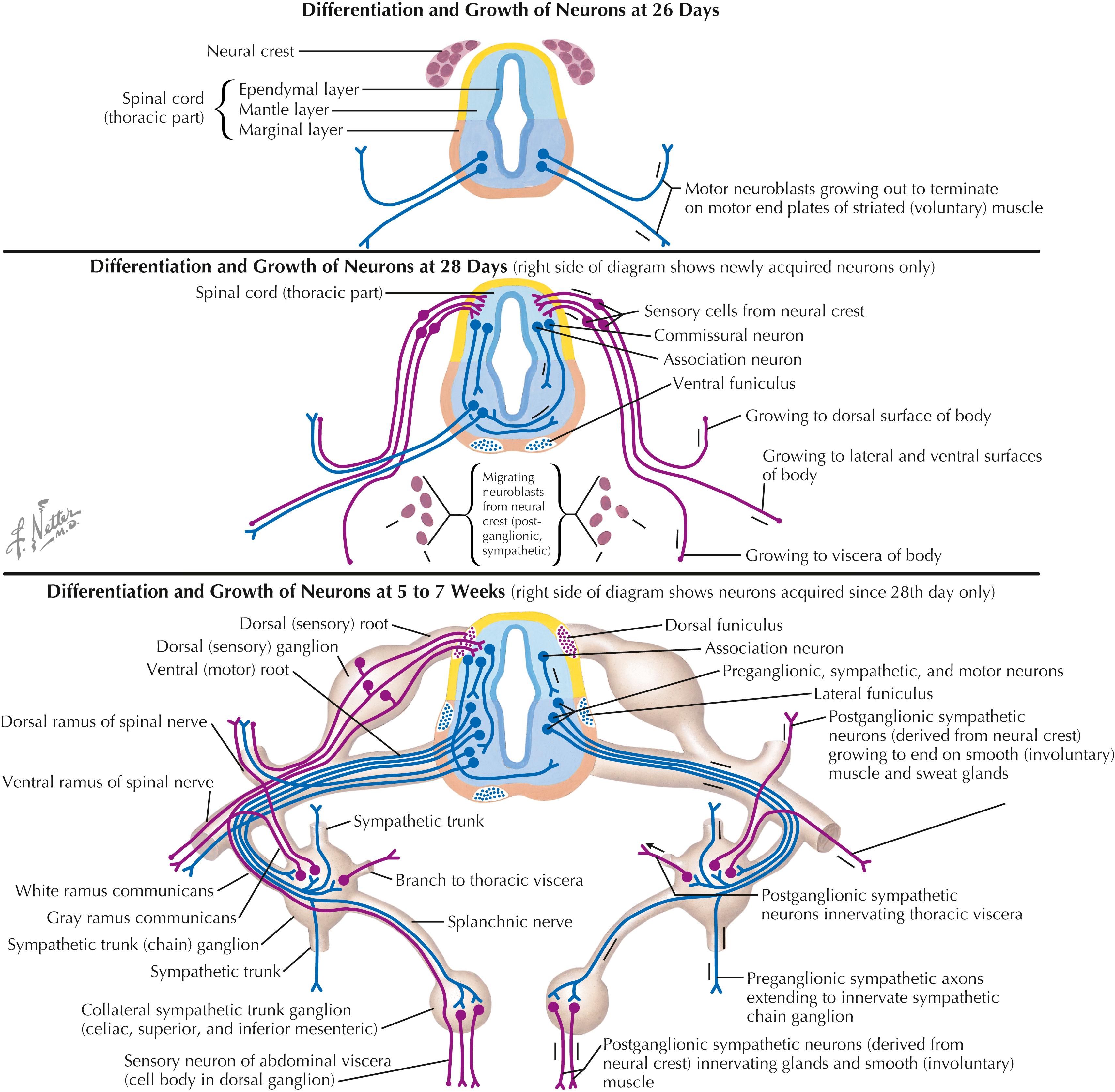
Peripheral axon development is a complex process of central and peripheral neurite extension, trophic and chemotactic factors, and axonal guidance and maintenance by innervated target tissues. Dorsal root ganglion cells are bipolar; a peripheral axonal process is associated with simple or complex sensory receptor cells, and a central axonal process extends into the central nervous system (CNS) to form connections with secondary sensory neurons. The lower motor neurons send motor axons to the developing skeletal muscles through the ventral roots or motor cranial nerves, forming neuromuscular junctions as sites of synaptic connectivity. Motor neurons that fail to establish such contact with skeletal muscles die. Central preganglionic axons exit in the ventral roots and terminate on sympathetic ganglion cells in the sympathetic chain or collateral ganglia or on parasympathetic intramural ganglia near the organs innervated. Postganglionic axons form connections with target tissues, including smooth muscle, cardiac muscle, secretory glands, some metabolic cells (hepatocytes, fat cells), and cells of the immune system in parenchymal zones of many lymphoid organs. Sensory, motor, and autonomic symptoms can occur in peripheral neuropathies based on disruption of these connections.

Somatopleure and splanchnopleure constitute the embryonic basis for the subdivision of the PNS into spinal (somatic) nerves and splanchnic (autonomic) nerves. The somatopleure develops from ectoderm and the somatic portion of lateral plate mesoderm. Somite hypoblasts migrate into somatopleure to form the lateral and ventral aspects of the body wall, including the limbs. Splanchnopleure, derived from endoderm and lateral plate mesoderm, give rise to visceral organs. The ventral rami migrate into somatopleure, and splanchnic nerves grow into splanchnopleure. Thoracic and lumbar splanchnic nerves have sympathetic and visceral sensory axonal components. Pelvic splanchnic nerves (S2–S4) have parasympathetic and visceral sensory axonal components.
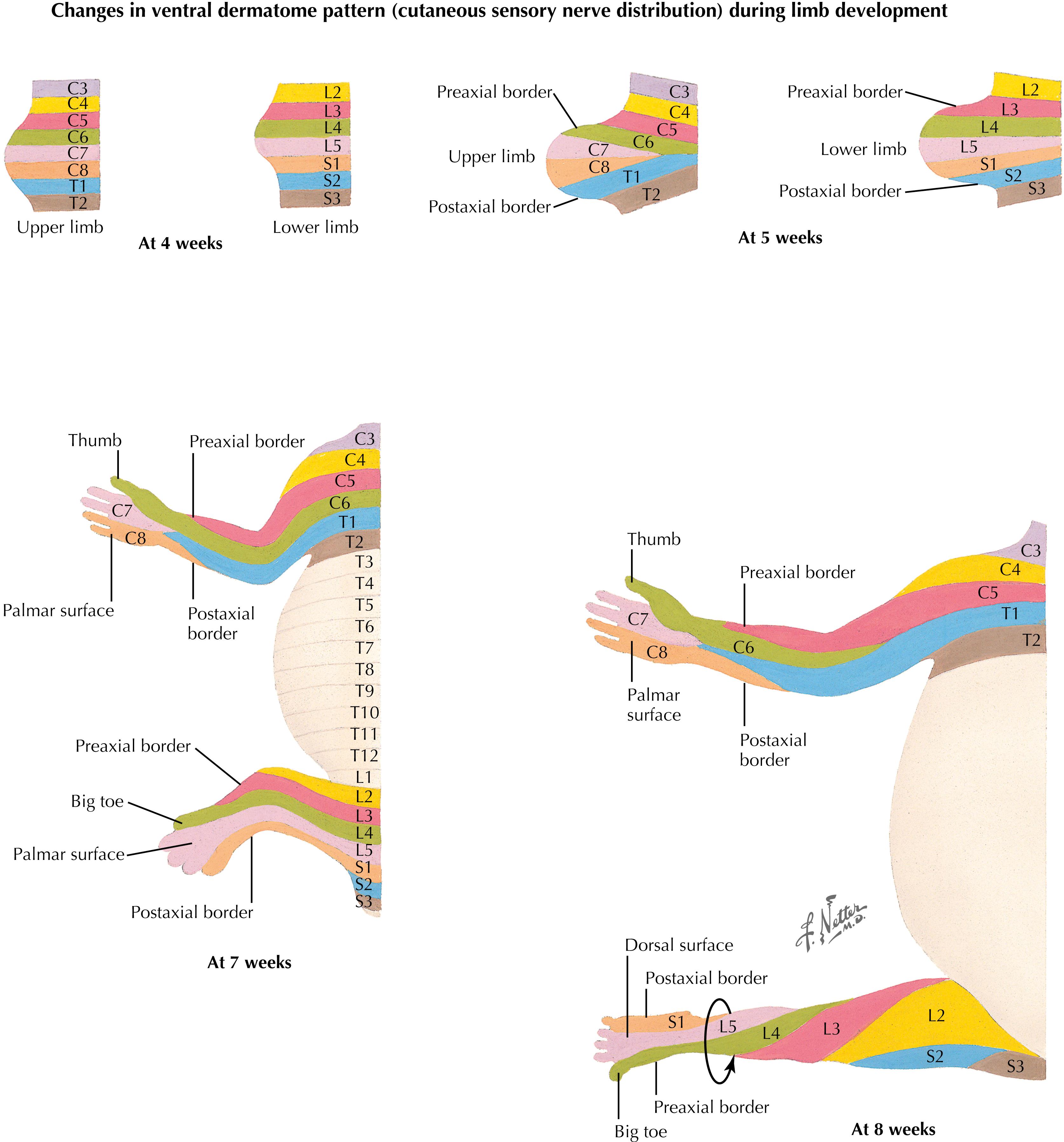
Rotation of the lower limb results in a reversal of the preaxial and postaxial borders, producing a spiral arrangement of dermatomes. Spinal nerve segments on the anterior surface of the lower extremity extend medially and inferiorly; the great toe (hallux) is supplied by nerves from a more rostral dermatome (L4) than the little toe (S1). The lower extremity is an extension of the trunk, and the most caudal dermatomes (sacral and coccygeal) supply the perineum, not the foot. Cervical dermatomes maintain a relatively orderly distribution to the upper extremity with minimal rotation.

Early in development (5 weeks), neuroblasts in the ependymal layer lining the central canal move back and forth from the ependymal surface to the pial surface, replicating as they go. Neural migration follows distinctive patterns in different regions of the neural tube. In the spinal cord, neurons migrate into the inner mantle zone, leaving the outer marginal zone as a site for axonal pathways. In the cerebellar cortex, some neurons migrate to an outer location on the outer pial surface as an external granular layer, from which granular cells then migrate inward to synapse with other neurons present in deeper layers of the cerebellar cortex. In the cerebral cortex, neurons migrate to the outer zone, where the gray matter (neuronal cell bodies) remains on the surface, external to the white matter (nerve fibers). These developmental patterns reflect the anatomical organization of the mature structures, their blood supply, and their vulnerability to injury by tumors, vascular insults, trauma, and other disorders.

Neural tube ependymal cells give rise to neuroblasts, from which the neurons of the CNS are derived. They also give rise to the glioblasts, from which the mature ependymal cells, astrocytes, and oligodendroglia are derived. Microglia, the “scavenger cells” of the CNS, are derived mainly from mesodermal precursors. Cells of glial origin are the predominant cells that give rise to CNS tumors. The neural crest cells give rise to many peripheral neural structures, including primary sensory neurons, postganglionic autonomic neurons of both the sympathetic and parasympathetic systems, adrenal medullary chromaffin cells, pial and arachnoid cells, Schwann cells (the supporting cells of the PNS), and some other specialized cell types. Neural crest cells can be damaged selectively in some disorders (e.g., familial dysautonomias) and also can give rise to specific tumor cell types such as pheochromocytomas. Most microglial cells are derived from specialized mesenchymal cells infiltrating from the yolk sac.
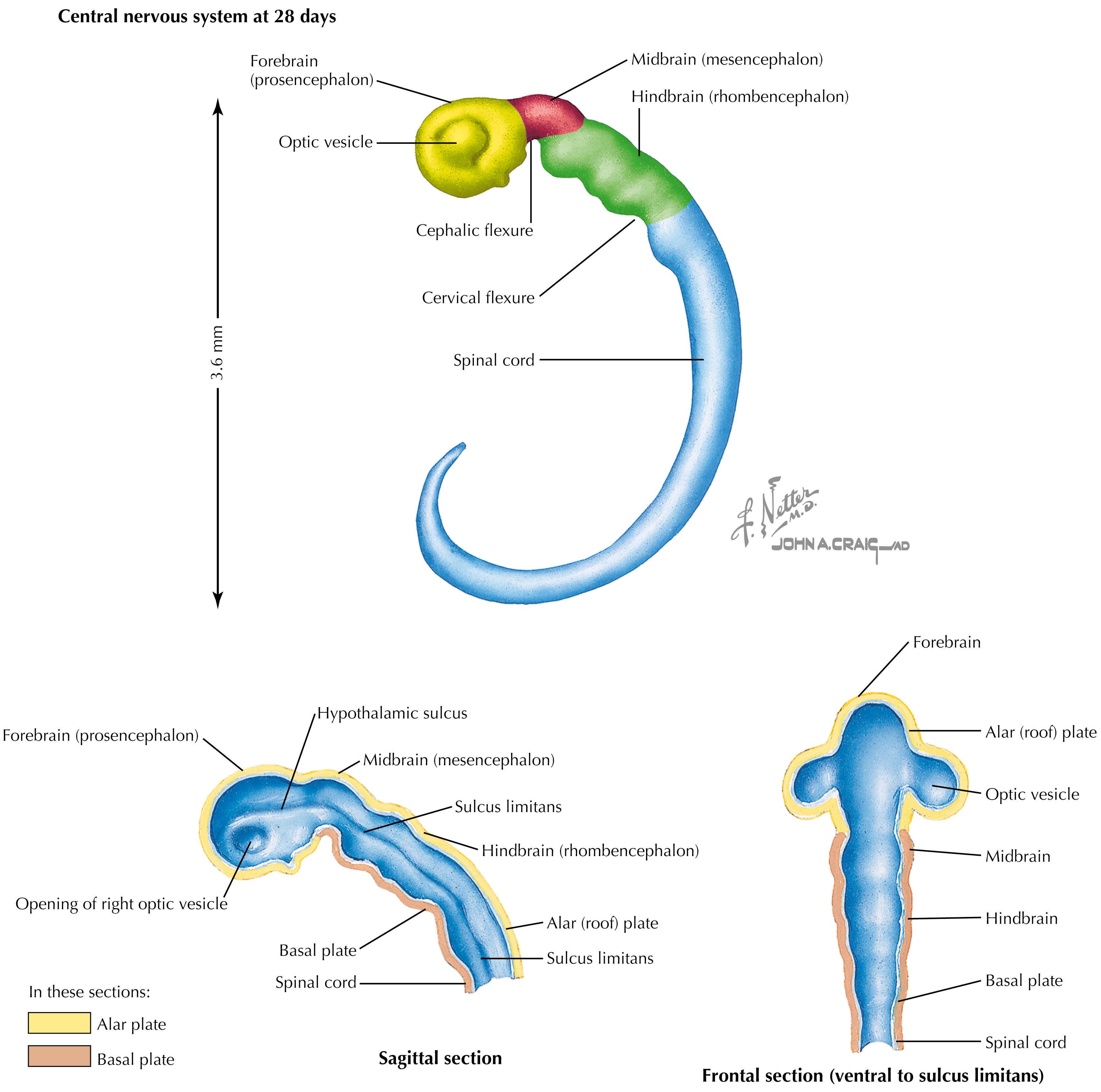
Some components of the neural tube expand differentially, resulting in bends or flexures that separate the neural tube into caudal to rostral components. The cervical flexure caudally and the cephalic flexure rostrally result from the differential expansion. Three regions of rapid cellular proliferation develop: the forebrain (prosencephalon) rostrally, the mesencephalon (midbrain) in the middle, and the hindbrain (rhombencephalon) caudally. The ventricular system bends and expands to accommodate the increasing neural growth. An outgrowth from the caudal part of the prosencephalon extends from the future diencephalon to become the optic cup, giving rise to the future retina and its central connections.
The optic vesicle develops from the prosencephalon, specifically the future diencephalon. As a consequence, the neuroretina is actually a central neural derivative and not a peripheral neural crest derivative. Therefore, the retina is supplied with CNS vasculature, and the ganglion cells of the retina (projecting into the optic nerve, chiasm, and tract) are actually CNS axons myelinated by oligodendroglia and surrounded by subarachnoid space and its cerebrospinal fluid. As a CNS tract, the optic nerve is subject to central demyelinating lesions as seen in multiple sclerosis. The retinal vasculature is the only CNS vasculature that is directly observable by ophthalmoscopy.

Become a Clinical Tree membership for Full access and enjoy Unlimited articles
If you are a member. Log in here