Physical Address
304 North Cardinal St.
Dorchester Center, MA 02124

See next page.
The autonomic nervous system is a two-neuron chain connecting preganglionic neurons through ganglia to visceral target tissues (cardiac muscle, smooth muscle, secretory glands, metabolic cells, cells of the immune system). The sympathetic division (sympathetic nervous system; SNS) is a thoracolumbar (T1-L2) system arising from the intermediolateral cell column of the lateral horn of the spinal cord, acting through chain ganglia and collateral ganglia; it is a system designed for enhancing activities and for fight-or-flight reactions in an emergency. The parasympathetic division (parasympathetic nervous system) is a craniosacral system arising from brainstem nuclei associated with cranial nerves (CNs) III, VII, IX, and X and from the intermediate gray in the S2–S4 spinal cord. Connections from CNs III, VII, and IX act through cranial nerve ganglia; connections from the vagal system and sacral system act through intramural ganglia in or near the target tissue. The parasympathetic nervous system is a homeostatic reparative system. Central connections from the limbic forebrain, hypothalamus, and brainstem regulating the sympathetic and parasympathetic nervous systems’ outflow to the body act mainly through connections to vagal and sympathetic preganglionic neurons.
Preganglionic parasympathetic neurons in the brainstem and sacral spinal cord, as well as preganglionic sympathetic neurons in the thoracolumbar spinal cord, send projections to ganglion cells and use acetylcholine as the principal neurotransmitter. The ganglion cells possess mainly nicotinic cholinergic receptors for transducing fast neurotransmission responses. Postganglionic sympathetic neurons use mainly norepinephrine as their neurotransmitter, whereas post-ganglionic parasympathetic neurons use acetylcholine. Target tissue possesses alpha and beta adrenoceptor subclasses and cholinergic muscarinic receptor subclasses (M1–M3). In the heart, beta1 receptors increase the force and rate of contraction, increase cardiac output, and dilate coronary arteries, whereas M2 receptors decrease the force and rate of contraction and cardiac output. In vascular smooth muscle and smooth muscles of the pupil, ureters, and bladder, alpha1 receptors cause contraction. In blood vessels, alpha2 receptors also cause constriction. In smooth muscle of the tracheobronchial system, uterus, and gastrointestinal tract vasculature, beta2 receptors cause relaxation. Alpha1 receptors cause relaxation of gastrointestinal smooth muscles, and M1 receptors cause slow contraction. M3 receptors cause contraction of most parasympathetic smooth muscle target structures. In salivary glands, alpha1 receptors cause secretion and beta2 receptors cause mucus secretion. In adipose tissue, alpha1 receptors cause glycogenolysis, beta1 receptors cause lipolysis, and alpha2 receptors inhibit lipolysis. In sweat glands, alpha1 receptors cause secretion. In the kidney, alpha1 receptors enhance reabsorption of Na + , and beta1 receptors provoke renin release. In liver and skeletal muscles, beta2 receptors cause glycogenolysis. In the pancreas, beta2 receptors stimulate insulin release, and alpha2 receptors inhibit insulin release. On immunocytes, beta-adrenergic receptors decrease natural killer (NK) cell activity and decrease the secretion of Th1 cytokines (interferon-gamma, interleukin 2) by Th1 lymphocytes. The balance of adrenergic and cholinergic neurotransmission determines the relative degree of activation of target tissues, and differential affinity of ligands for the various receptor subclasses helps to determine the final integrative physiological response.
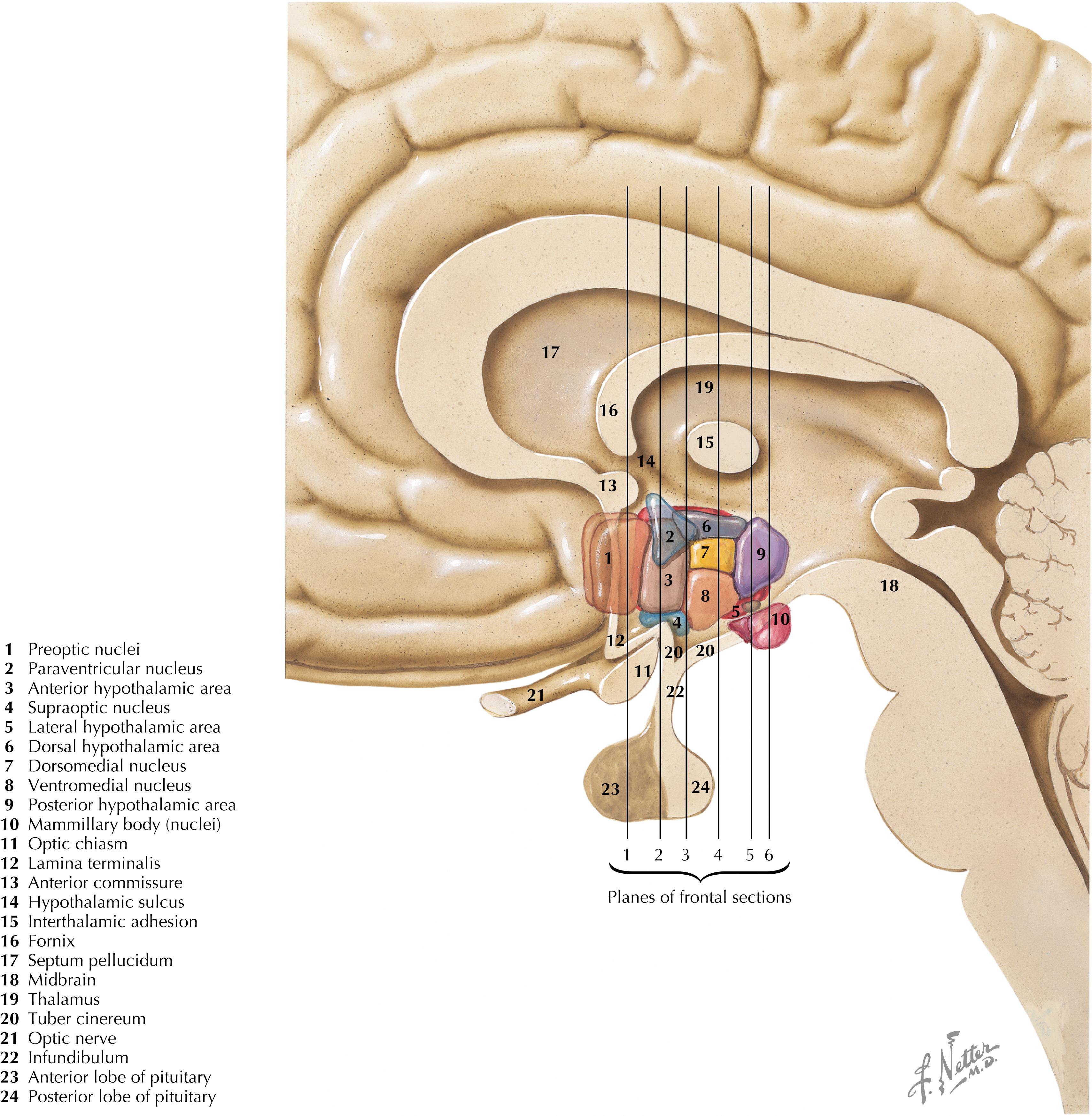
The hypothalamus is a collection of nuclei and fiber tracts in the ventral diencephalon that regulates visceral autonomic functions and neuroendocrine functions, particularly from the anterior and posterior pituitary. Many nuclei are found between the posterior boundary (mammillary bodies) and the anterior boundary (lamina terminalis, anterior commissure) of the hypothalamus; these nuclei are subdivided into four general hypothalamic zones, from rostral to caudal: (1) preoptic, (2) anterior or supraoptic, (3) tuberal, and (4) mammillary or posterior. From the medial boundary at the III ventricle to the lateral boundary, the nuclei are subdivided into three general zones or areas: (1) periventricular, (2) medial, and (3) lateral. The pituitary gland is attached at the base of the hypothalamus by the infundibulum (pituitary stalk), which possesses an important zone of neuroendocrine transduction, the median eminence.
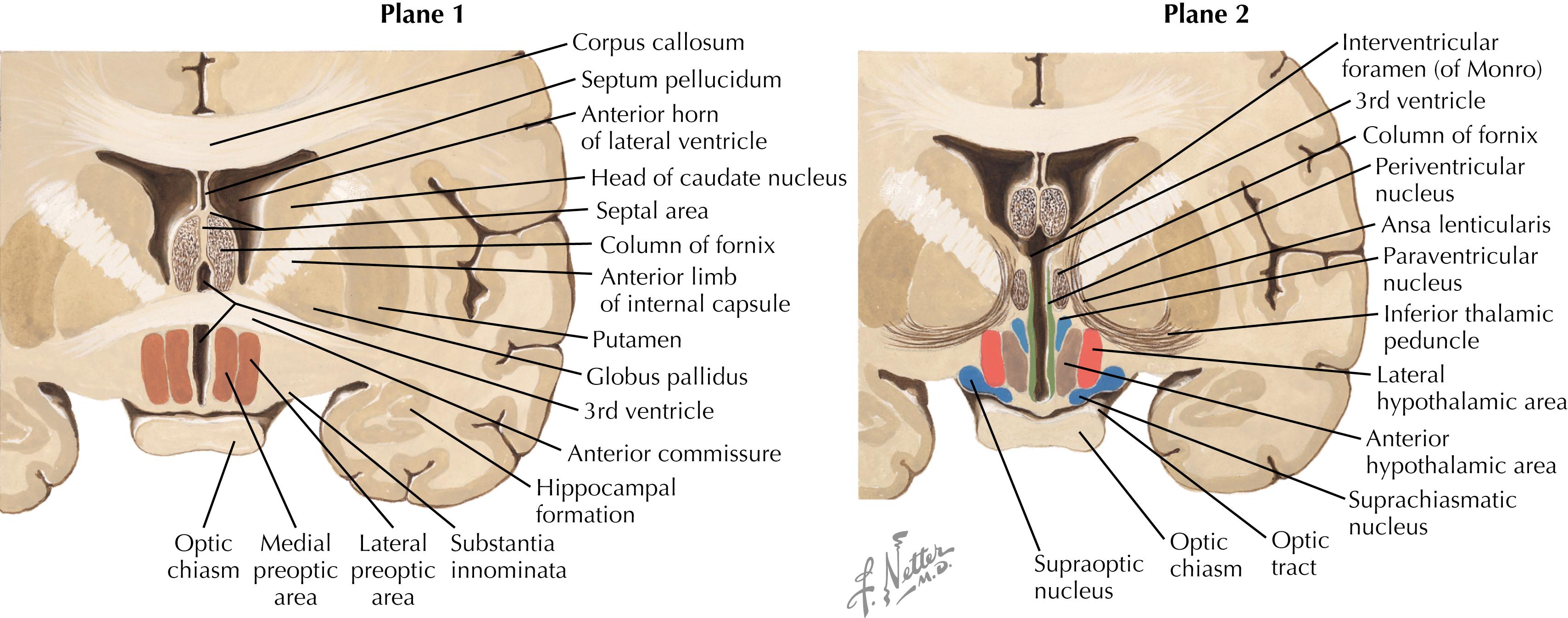
The major nuclei in the preoptic zone (Plane 1) include the medial and lateral preoptic areas. The organum vasculosum of the lamina terminalis (OVLT), a circumventricular organ (with no blood-brain barrier), is present in this hypothalamic area. The major nuclei in the supraoptic (anterior) zone (Plane 2) include the supraoptic (SON) and paraventricular (PVN) nuclei, the suprachiasmatic nucleus, the anterior hypothalamic area, and the lateral hypothalamic area (LHA). Some nuclei such as the PVN have many subregions (such as the magnocellular and parvocellular regions) that contain many collections of chemically specific neurons (20 or more) that have discrete projections and functions. These groups are sometimes intermingled within one subregion of the nucleus.
The hypothalamus and brainstem structures are involved in regulating the sleep-wake cycle . Ablative lesions of the preoptic area result in insomnia. Some preoptic neurons appear to be maximally activated during sleep and may inhibit neurons in the posterior hypothalamus (such as tuberomammillary neurons) that contribute to wakefulness. The LHA also contains neurons involved in wakefulness through the secretion of an activating neuropeptide, hypocretin. Neurons of the LHA activate the tuberomammillary neurons as well as the locus coeruleus in the pons, a noradrenergic cell group with widespread projections to all regions of the central nervous system (CNS) and a major role in arousal and wakefulness. Early epidemics of encephalitis lethargica (sleeping sickness) demonstrated damage to the midbrain and posterior regions of the hypothalamus. This scheme is consistent with a role for the posterior hypothalamus in sympathetic activation and arousal and with a role for the anterior and preoptic hypothalamus in parasympathetic activation and quiet, reparative, homeostatic functions. Narcolepsy is a condition of episodic periods of overwhelming daytime drowsiness and then an abrupt episode of sleep, even in the middle of an activity. The person then awakens and feels alert. Nighttime sleep may be disturbed, but this is not the cause of daytime sleep episodes; patients with narcolepsy go into rapid eye movement sleep in a matter of minutes rather than hours. Many stimuli (e.g., intense emotion, excitation, laughter) may precipitate an episode of cataplexy in which the knees give out, the person falls, and an abrupt sleep episode follows. Sleep apnea is a major sleep disorder, often associated with obesity, in which patients have prolonged periods of apnea, followed by gasping and including disturbed sleep and loud snoring. It is a major risk factor for heart disease.
The suprachiasmatic nucleus (SCN) sits just above the optic chiasm and contains the major neurons of the CNS that act as a “pacemaker” system for the control of diurnal, or circadian, rhythms . The intrinsic pacemaker has a cycle that is a bit longer than 24 hours (studied in humans who lived in caves with no external light cues); however, input from the retina to the suprachiasmatic nucleus entrains the diurnal rhythms to a 24-hour period. These diurnal rhythms drive many hormone and metabolic levels (e.g., cortisol is low in the late evening and high in the morning before rising; melatonin is highest in late evening) and physiological functions (blood pressure and core body temperature are lowest in early morning and highest in late afternoon). Superimposed on these diurnal rhythms are broader factors, such as effects of the sleep-wake cycle, life stress, levels of activity, and other environmental factors. Sleep has a particularly important influence on cortisol rhythms. Disrupted or poor sleep habits can ablate the diurnal cortisol rhythm, leading to a propensity for fat to be deposited in a central abdominal location because of the effects of high cortisol levels. This can contribute to the likelihood of metabolic syndrome , with its elevated inflammatory mediators (C-reactive protein and interleukin [IL]-6) and increased risk for cardiovascular disease, stroke, type II diabetes, and many cancers. The SCN is influenced by a host of limbic and other forebrain influences superimposed on diurnal rhythms. The SCN, in turn, has axonal projections to other regions of the hypothalamus, the locus coeruleus, and limbic sites through which the diurnal regulatory control of these hormones and physiological functions is achieved.
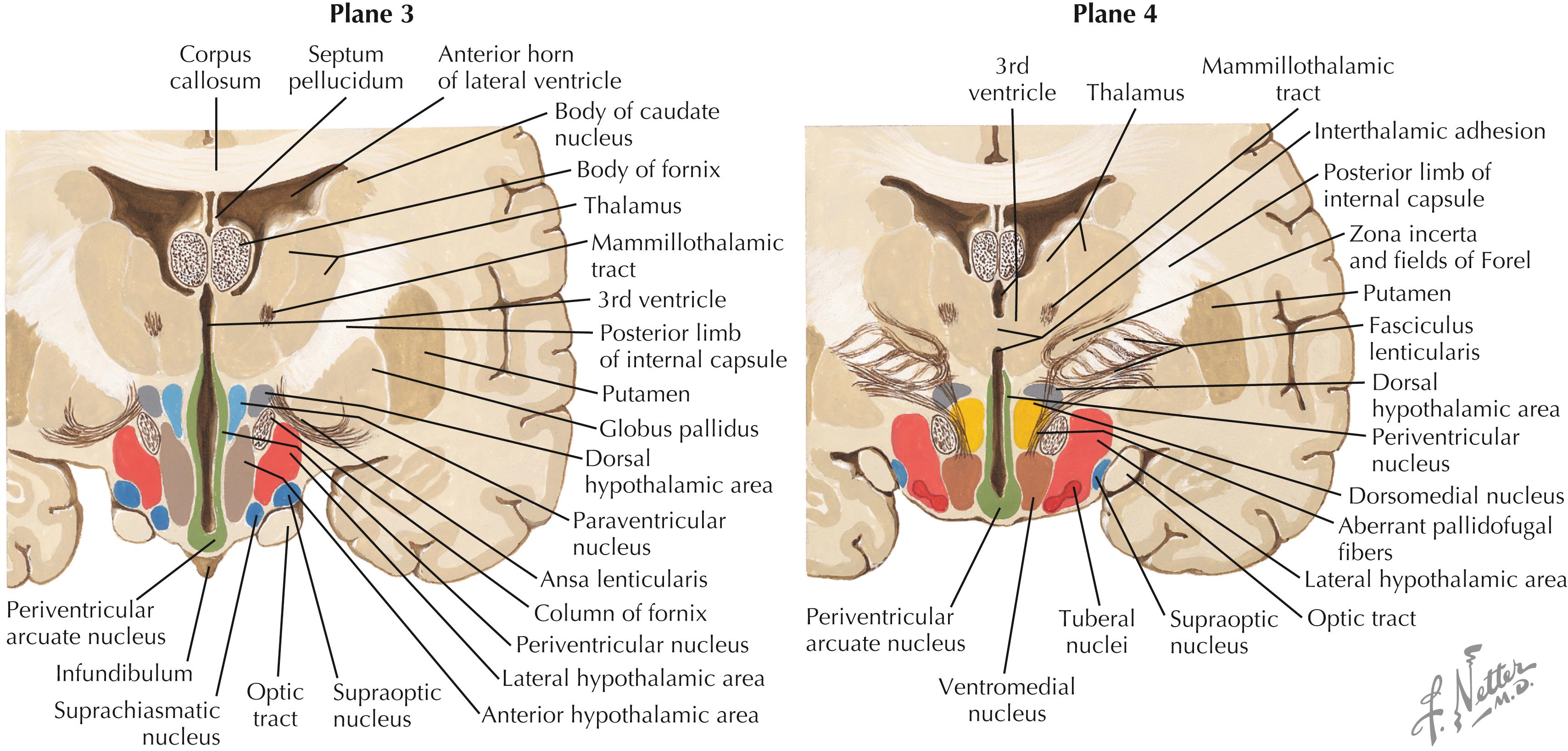
The major nuclei in the tuberal zone (Planes 3 and 4) include the dorsomedial nucleus, the ventromedial nucleus, the periventricular area or nucleus, the arcuate nucleus, the periarcuate area (beta-endorphin cells), the tuberal nuclei, the dorsal hypothalamic area, and the LHA. Some nuclei from the supraoptic zone (PVN, SON, LHA) extend caudally into this zone. The median eminence extends from this region, and axons from releasing-factor and inhibitory-factor neurons that control the release of anterior pituitary hormone funnel down to the contact zone, where they release these factors (hormones) into the hypophyseal portal system, which bathes the cells of the anterior pituitary.
The secretion of hormones by the anterior pituitary gland is regulated by releasing factors (hormones) and inhibitory factors (hormones) that are produced by neurons of the hypothalamus and adjacent sites and are secreted by their axons into the hypophyseal portal vasculature for delivery in extraordinarily high concentrations to cells of the anterior pituitary. A well-known releasing factor is corticotropin–releasing hormone or factor (CRH or CRF), produced by parvocellular neurons of the paraventricular nucleus, which regulates subsequent secretion of adrenocorticotropic hormone (ACTH) and cortisol. Another important releasing hormone, growth hormone-releasing hormone, is produced by neurons in the arcuate nucleus and delivered by their axons to the hypophyseal portal system. Somatostatin is a growth hormone-inhibitory hormone and is produced by other neurons in the arcuate nucleus as well as elsewhere. These hormones are regulated by neural connections, hormonal influences, and metabolic factors.
Growth hormone (GH) is released in pulsatile bursts during stage 3 and stage 4 sleep, accounting for 70% of GH release. GH release also is stimulated by exercise, acute stressors, hypoglycemia, and intake of protein, and is suppressed by intake of glucose and many fatty acids. Children who experience emotional deprivation secrete low levels of GH and may fail to grow. Recent studies have shown that mirthful laughter associated with viewing humorous videos markedly stimulates GH secretion and diminishes cortisol and epinephrine secretion. Even more remarkable, when subjects anticipate viewing something humorous, the anticipation itself provokes GH secretion that is as great as or greater than the GH secretion seen in stage 3 and stage 4 sleep.
Sex steroid hormones influence brain development. In a male fetus, the developing testes provide androgens (converted in the brain to estradiol) that influence CNS development in a male pattern during critical developmental periods. All developing fetuses are exposed to maternal estrogen as well as some placental hormones, but the estrogen is bound by alpha-fetoprotein, which protects the female fetus from masculinization by the CNS. One important consequence of fetal exposure to sex steroids is the subsequent hypothalamic control of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) from the anterior pituitary gland. In females, these hormones are released in a cyclic fashion. In males, FSH and LH are released in steady amounts, a phenomenon dependent upon CNS exposure to estradiol via androgens during fetal development. In the CNS, FSH and LH secretion is controlled by gonadotropin-releasing hormone (GnRH), formerly called luteinizing hormone-releasing hormone. GnRH neurons in the preoptic area project to the contact zone of the median eminence, ending on the hypophyseal-portal vessels. The GnRH neurons are responsive to estrogen in the female brain but not in the male brain, perhaps accounting for the cyclic secretion of FSH and LH in females. The ventromedial (VM) nucleus of the hypothalamus appears to control some aspects of sexual behavior; VM neurons respond to progesterone via receptors in the female brain but not in the male brain. The male brain responds behaviorally to circulating androgens but not to estrogen. Anatomically, preoptic and VM neurons show male-female differences in morphological and synaptic features. A specialized portion of the preoptic area, the sexually dimorphic nucleus, is considerably larger in the male brain than in the female brain, apparently triggered by developmental hormonal exposure.
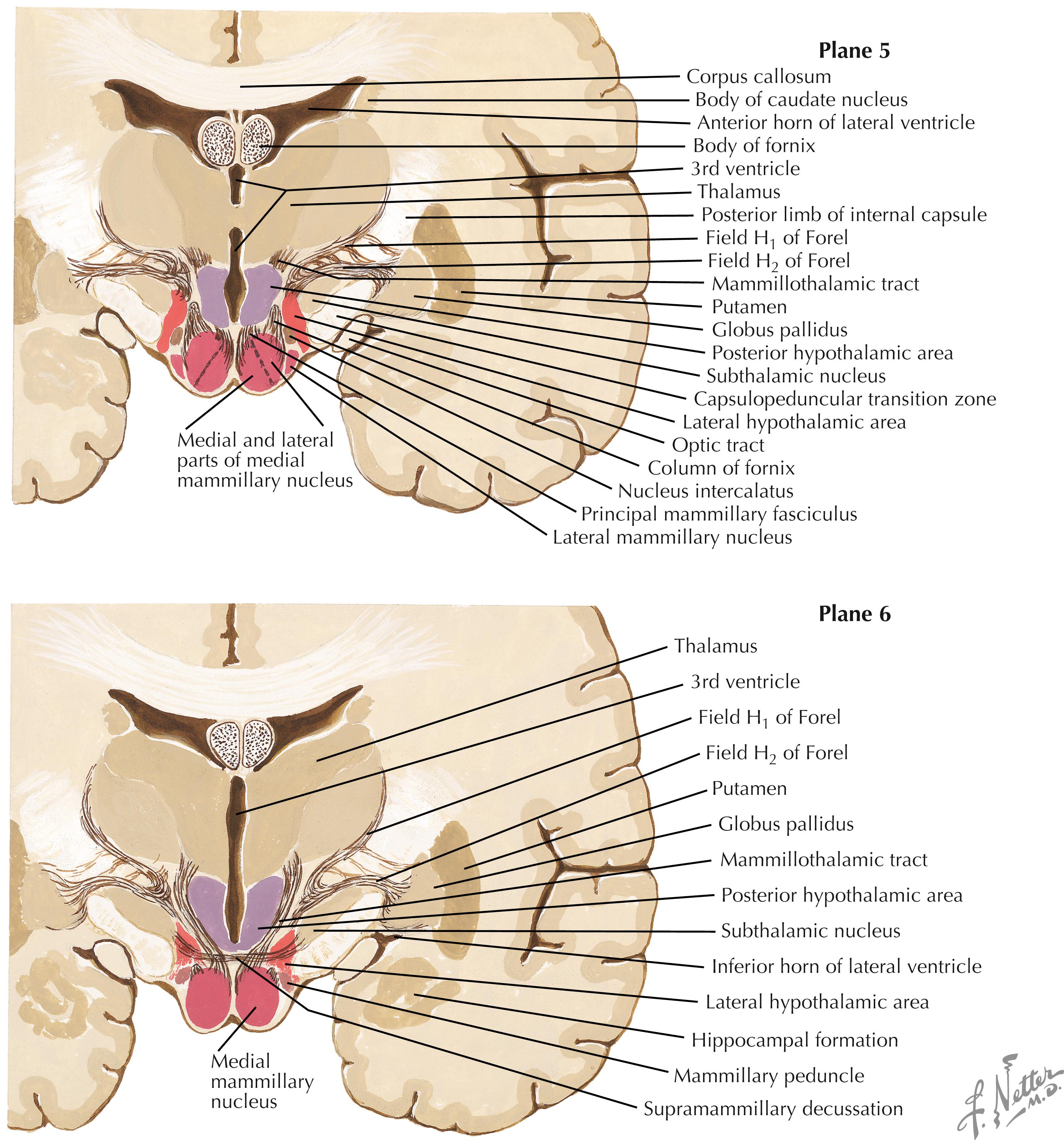
The major nuclei in the mammillary zone (Planes 5 and 6) include the medial and lateral mammillary nuclei, the posterior hypothalamic area, and the LHA. The LHA extends throughout most of the length of the hypothalamus and shows neuronal characteristics seen in the brainstem reticular formation.
In the 1930s, James Papez proposed a brain circuit that was viewed as a substrate for control of emotional behavior and later as a substrate for memory, especially for consolidation of immediate and short-term memory into long-term memory. This Papez circuit includes hippocampal formation (especially the subiculum) via the fornix to the mammillary nuclei (especially medial nuclei); via the mammillothalamic tract to the anterior thalamic nuclei; via the internal capsule to the anterior cingulate cortex; and via polysynaptic connections in the cingulum to the entorhinal cortex, subiculum, and hippocampus. This circuit is proposed as a site of major damage in Wernicke-Korsakoff syndrome , a disorder that is commonly seen in chronic alcoholic patients with a vitamin B 1 (thiamine) deficiency. This syndrome includes Wernicke’s encephalopathy and the memory dysfunction of Korsakoff’s syndrome. Wernicke’s encephalopathy involves a confused and psychotic state involving confabulation (made-up stories derived from a host of confused past memories or experiences), cerebellar ataxia, extraocular and gaze palsies, and nystagmus. Korsakoff amnestic syndrome involves the inability to consolidate immediate and short-term memory into long-term traces (anterograde amnesia) as well as long-term memory loss concerning events that have occurred since the onset of the disease. Degeneration has been described in the mammillary bodies, fornix, hippocampal formation, and anterior and medial dorsal thalamus. However, the extent to which the mammillary nuclei themselves play a role in consolidation of memory traces remains to be shown. Thiamine administration may help to reverse some of the symptoms, but the amnesias may persist. Administration of glucose (carbohydrate loading) without thiamine may cause death as the result of nutritional cardiomyopathy.
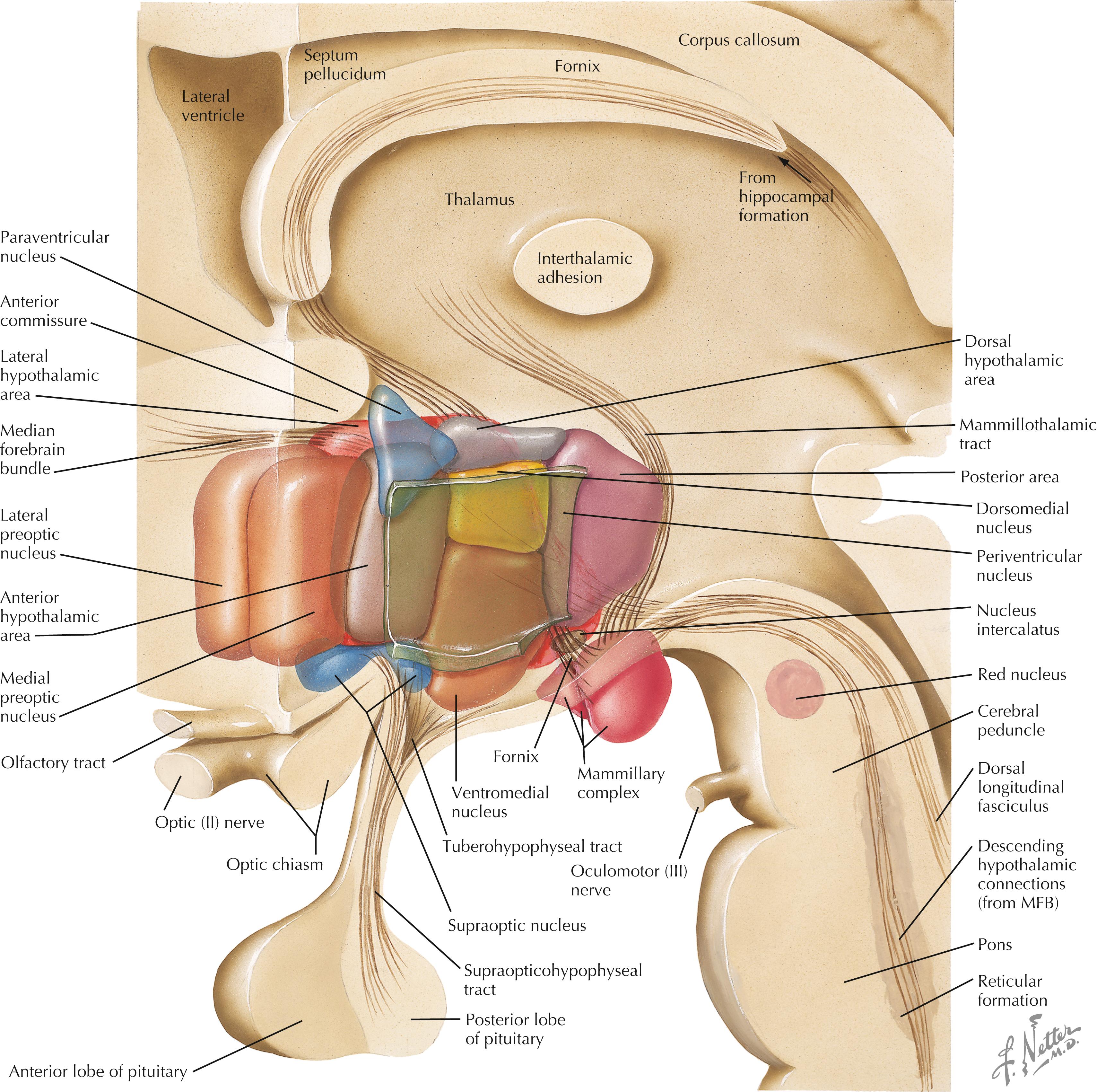
A schematic three-dimensional reconstruction of the hypothalamus in sagittal section shows the nuclei, areas, and zones that occupy this small, compact region of the diencephalon. Many pathways are represented in this schematic reconstruction, including the fornix, the mammillothalamic tract, the median forebrain bundle (MFB), the supraopticohypophyseal tract, the tuberohypophyseal (tuberoinfundibular) tract, and brainstem connections with the hypothalamus via the dorsal longitudinal fasciculus, the descending median forebrain bundle, the mammillotegmental tract, and descending connections from the PVN to preganglionic autonomic nuclei.
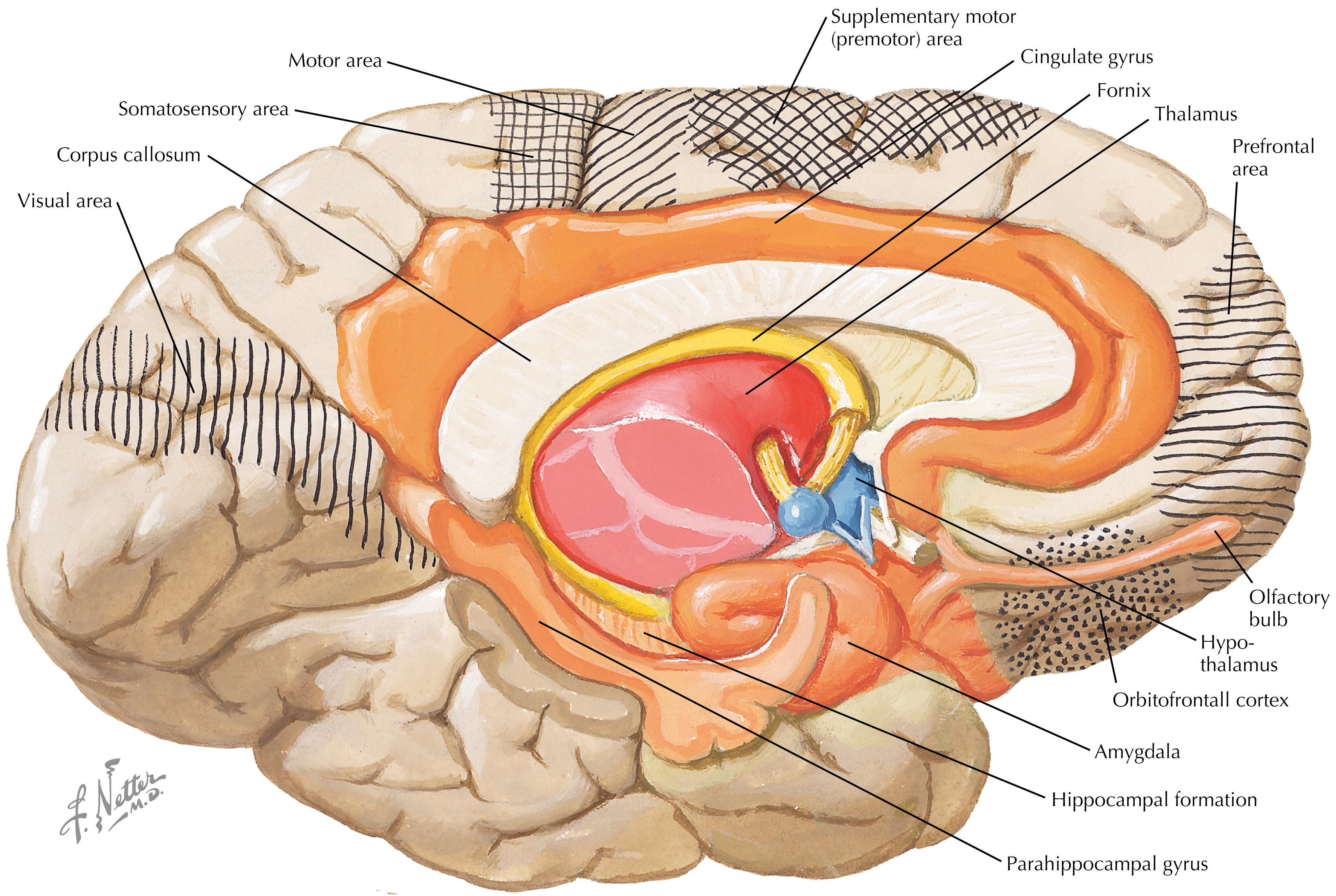
Numerous forebrain regions are intimately connected with the hypothalamus, some through direct fiber projections and others through indirect connections. These important regions of the cerebral cortex include the prefrontal cortex, orbitofrontal cortex, cingulate cortex, insular cortex, parahippocampal cortex, and periamygdaloid cortex. The important subcortical regions of the limbic forebrain include the hippocampal formation (a three-layer archicortex), amygdaloid nuclei, and septal nuclei. Important thalamic connections include the medial dorsal and anterior nuclei. Important olfactory connections include the olfactory tract, nuclei, and cortex.
The placebo effect is a positive change in a patient’s symptoms or subjective experience, or in a patient’s physiological state, including pain modulation, altered cardiovascular function, and immune reactivity (both innate and acquired), based on the patient’s expectations, interactions with health care personnel and treatments, or administration of medication (e.g., a pill) that normally has little direct pharmacological effects. Negative effects from such expectations or interactions are called a nocebo effect. The placebo effect has been described as “not real, ” “not a medicine,” “based on belief,” and having “no effect on disease processes or illnesses.” However, in pharmaceutical testing of highly reactive medications, it is often observed that the placebo is almost as effective as the pharmacological medication for altering clinical outcomes. Placebo effects, often involving conditioned responses, do indeed alter physiological processes, sometimes profoundly, with effects on disease outcome, as happens with conditioned immune responses in which a “placebo” alters lethal outcomes in experimental models of immune-related diseases (see the work of Robert Ader and Nicholas Cohen).
Placebo effects occur through known pathways and circuitry of the brain, including prefrontal cortex, anterior insular cortex, rostral anterior cingulate cortex, some amygdaloid nuclei, and in the brainstem, the periaqueductal gray. These structures act through influences on autonomic and neuroendocrine outflow, as well as by initiating appropriate behavioral responses. Disruption of these forebrain circuits can prevent the physiological effects of the placebo. Neurotransmitter systems such as endorphins, cannabinoids, dopamine and other catecholamines, and cortisol are involved in mediating placebo effects, and their pharmacological alterations (e.g., naloxone blockade of opioid receptors in placebo administration for pain) can prevent the physiological and behavioral alterations seen in the placebo effect.
Use of placebo effects and acknowledgment of conditioned responses have an important role in clinical medicine and disease treatment. It is likely that many complementary medicine approaches utilize, in part, placebo effects with their appropriate forebrain circuitry and neurotransmitter systems. This is consistent with the “relaxation response” described and documented by Herb Benson and colleagues, and the use of guided imagery, meditation, qi gong, and other parasympathetic-inducing practices.
See references for thoughtful discussions of placebo effect:
Colloca L, Barsky AJ: Placebo and nocebo effects. N Engl J Med 382:555–562, 2020.
Finnias DG, Kaptchuk TJ, Miller FG, Bennetti F: Biological, clinical, and ethical advances of placebo effect. Lancet 375:686-695, 2010.
Kaptchuk TJ, Miller FG: Placebo effects in medicine. N Engl J Med 373:8-9, 2015.
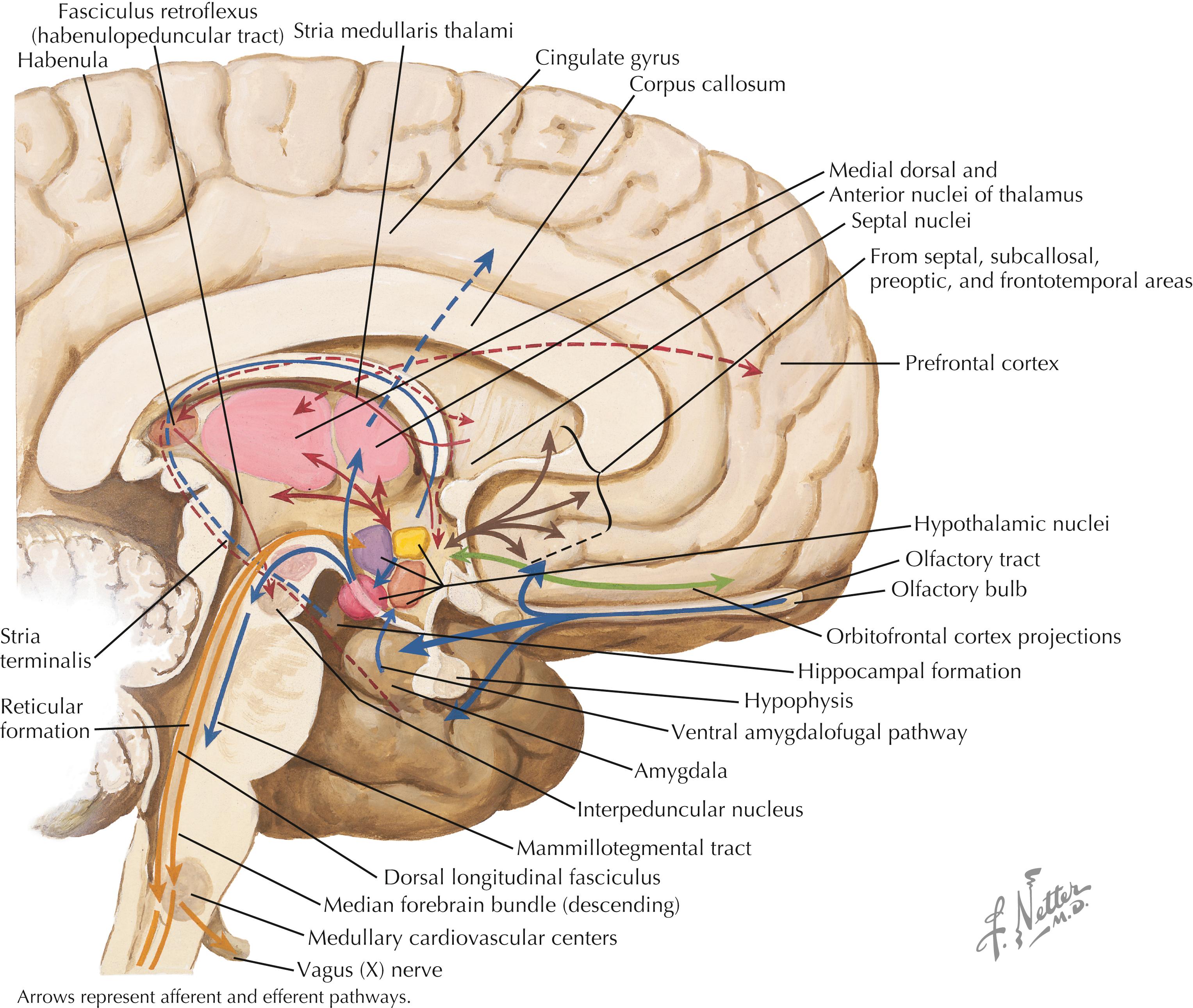
Hypothalamic connections are numerous and complex. Some regions of the cerebral cortex (prefrontal, orbitofrontal) and thalamus (anterior) send axonal projections directly to the hypothalamus. Diverse afferent pathways arise from the hippocampal formation and the subiculum (fornix), amygdaloid nuclei (stria terminalis, ventral amygdalofugal pathway), and habenula (fasciculus retroflexus). The retina sends direct retinohypothalamic fibers to the suprachiasmatic nucleus of the hypothalamus. Numerous brainstem projections, some compact and some diffuse, ascend to the hypothalamus by multiple pathways (not shown here). Efferent connections from the hypothalamus include those to the median eminence (from multiple nuclei), the posterior pituitary (supraopticohypophyseal tract), the septal nuclei and the anterior perforated substance (median forebrain bundle), the thalamus (mammillothalamic tract), and many brainstem and spinal cord sites (dorsal longitudinal fasciculus, median forebrain bundle, mammillotegmental tract, direct connections from PVN to preganglionic neurons, and others). The habenula receives afferents from the septal nuclei, lateral preoptic hypothalamic region, and anterior thalamic nucleus via the stria medullaris thalami, and sends projections to the preoptic area and septal nuclei.
The hypothalamus receives inputs from the hippocampal formation and subiculum, amygdaloid nuclei, habenula, retina, some cortical areas, and many brainstem regions; a good number of these inputs are limbic forebrain and brainstem connections. The role of the hypothalamus is to regulate the visceral milieu and neuroendocrine secretion, particularly via the anterior and posterior pituitary. The efferents of the hypothalamus reflect this role and are directed to the posterior pituitary and contact zone of the median eminence (for control of anterior pituitary hormonal secretion), some limbic forebrain structures, and widespread areas of the brainstem and spinal cord that are involved in autonomic and visceral regulation. These connections help to coordinate appropriate behavioral responses to external and internal inputs and perceived challenges in the environment. The posterior and lateral hypothalamic regions are particularly involved in sympathetic drive and activational responses, such as the acquisition of food and water, the increase of core body temperature, sympathetic arousal, activities involved in aggressive interactions with the environment, and wakefulness states. Many of these activities are coordinated through connections in the median forebrain bundle. In contrast, the anterior and medial hypothalamic regions are particularly involved in parasympathetic functions, such as satiation, decreased core body temperature, quiet and reparative homeostatis-related activities, and sleep. Many of these activities are coordinated through connections in the dorsal longitudinal fasciculus and other descending pathways.
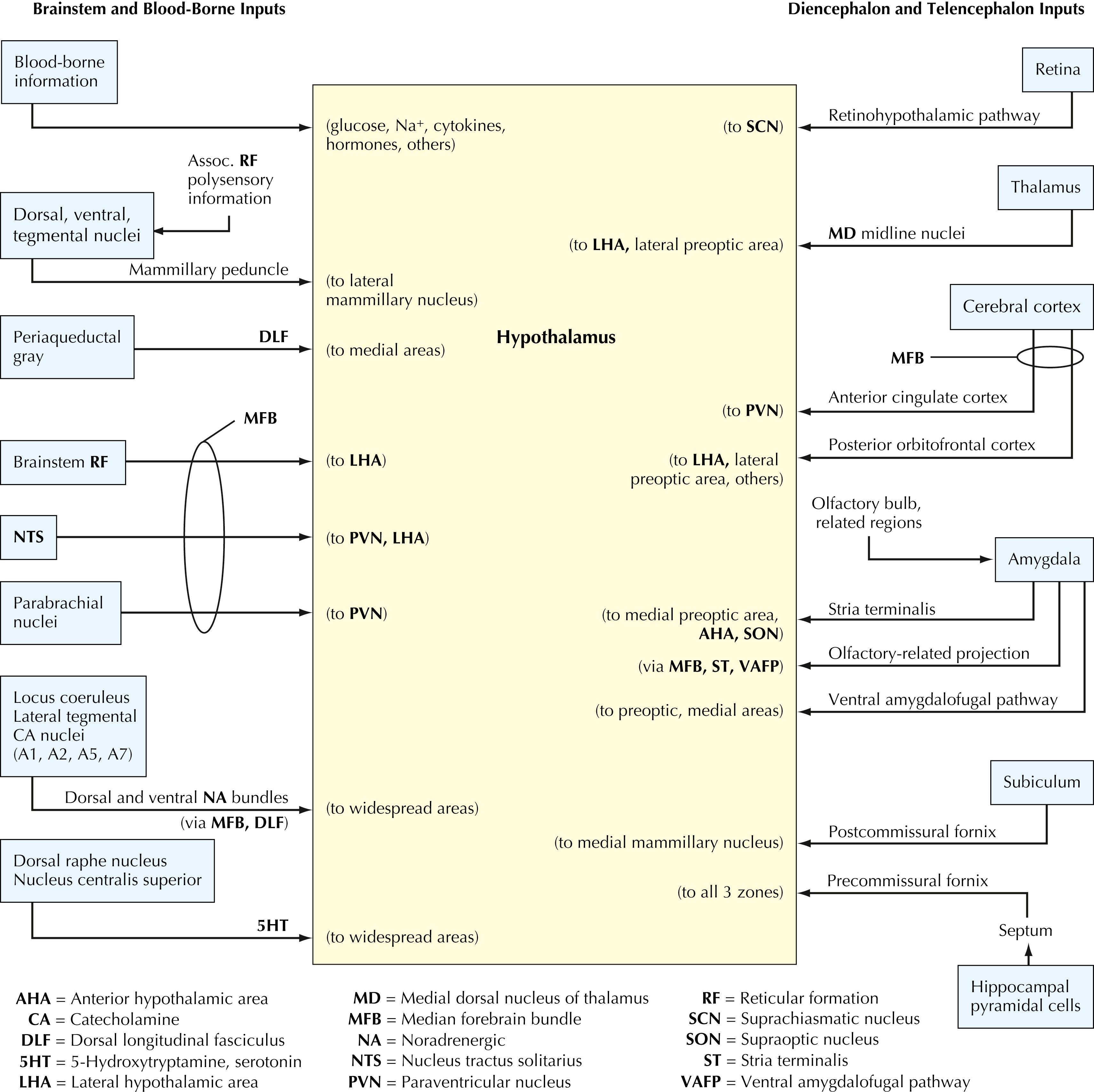
The hypothalamus receives extensive input from many regions of the CNS. Descending inputs arrive from limbic forebrain structures (hippocampal formation, subiculum, amygdaloid nuclei), the cerebral cortex (anterior cingulate, orbitofrontal, prefrontal), and the thalamus (medial dorsal). Ascending inputs arrive from extensive areas of the autonomic brainstem (tegmental nuclei, periaqueductal gray, parabrachial nuclei, nucleus tractus solitarius, locus coeruleus and tegmental catecholamine nuclei, raphe serotonergic nuclei) and from the brainstem reticular formation. The retina sends input directly to the suprachiasmatic nucleus, a nucleus of the hypothalamus that modulates diurnal rhythms. Blood-borne substances (cytokines, hormones, glucose, Na + , others) influence the hypothalamus via numerous routes and mechanisms.
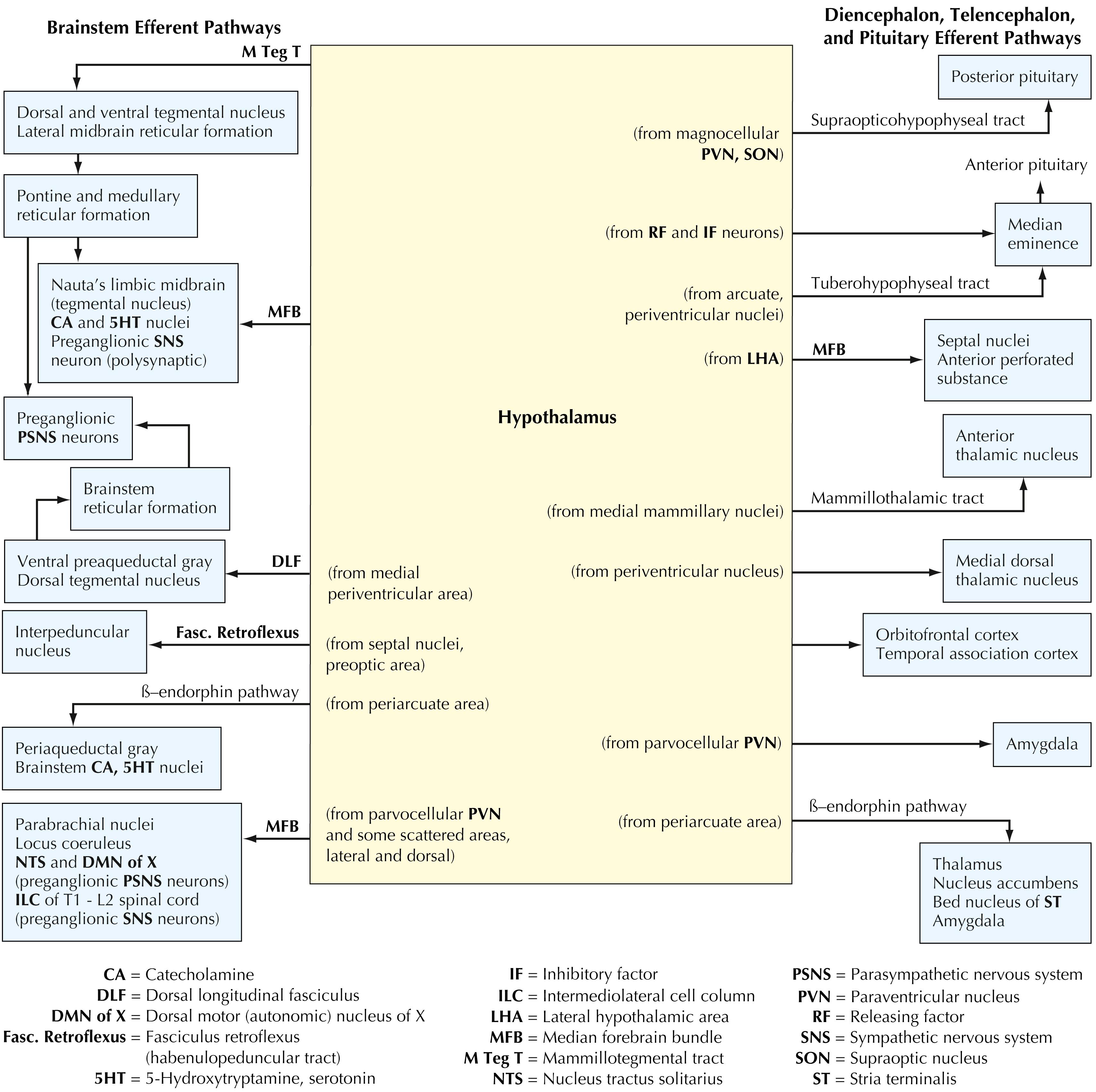
The hypothalamus gives rise to extensive efferent projections to many regions of the CNS. Ascending efferents are sent to limbic forebrain structures (amygdaloid nuclei, septal nuclei, the anterior perforated substance), the cerebral cortex (orbito frontal cortex and temporal association cortex), and the thalamus (medial dorsal, anterior). Extensive projections are sent to the median eminence (releasing and inhibitory factors for control of anterior pituitary hormones, dopamine projections from the arcuate nucleus and periventricular nucleus) and to the posterior pituitary. Additional efferent projections are sent directly and indirectly to the preganglionic neurons of the sympathetic and the parasympathetic nervous systems (median forebrain bundle, dorsal longitudinal fasciculus, mammillotegmental tract, and direct projections from the paraventricular nucleus), to widespread autonomic and visceral nuclei (noradrenergic neurons, serotonergic neurons, parabrachial nuclei, nucleus tractus solitarius, periaqueductal gray, tegmental nuclei, interpeduncular nucleus), and to the brainstem reticular formation.
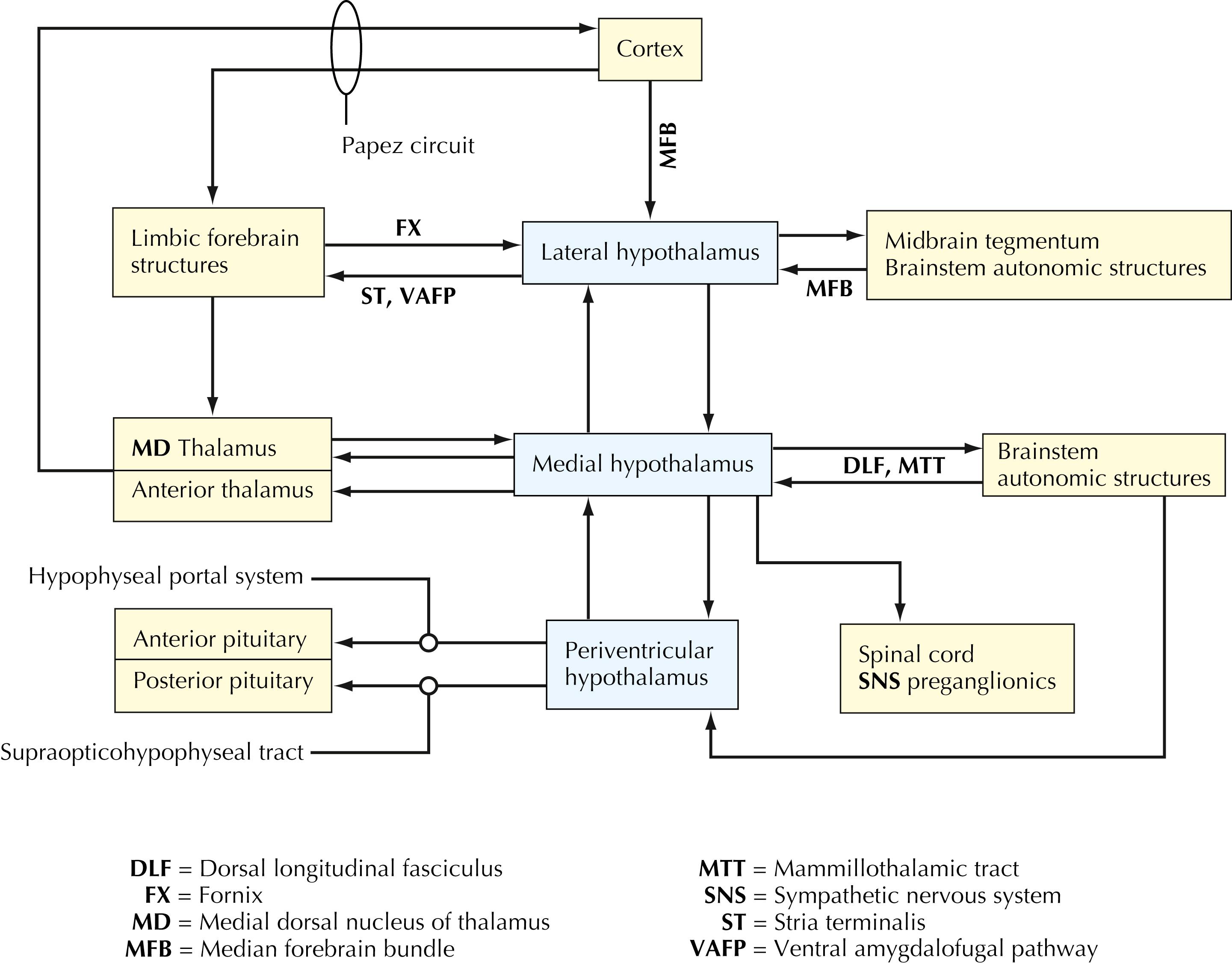
The lateral, medial, and periventricular zones of the hypothalamus have specific connections with the cerebral cortex, limbic forebrain structures, thalamus, and widespread areas of the brainstem. Extensive efferent projections of the hypothalamus are directed toward regulation of preganglionic sympathetic and parasympathetic neurons and toward release and regulation of hormones of the anterior and posterior pituitary. The anterior pituitary hormones regulate hormonal secretion and functional activities of many target structures throughout the body.
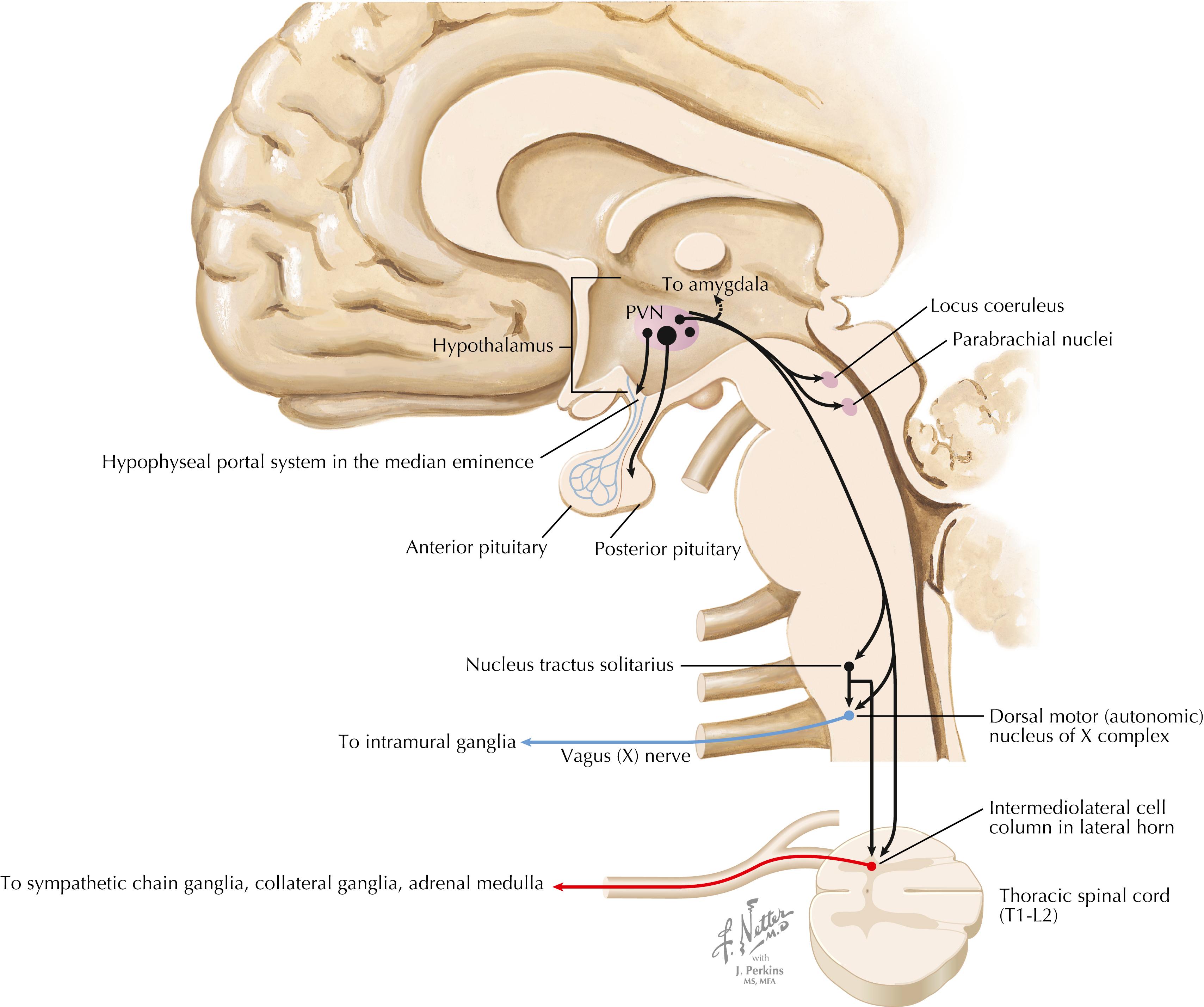
The PVN has many projections that help to coordinate pituitary neurohormonal outflow, autonomic preganglionic outflow, and limbic activity. Magnocellular neurons send axons to the posterior pituitary, releasing oxytocin and vasopressin into the general circulation. Corticotropin-releasing factor (CRF) neurons and some vasopressin neurons send axons to the median eminence; these axons release their hormones into the hypophyseal portal system, influencing the release of ACTH. PVN parvocellular neurons send direct descending projections to preganglionic neurons for the parasympathetics (dorsal motor nucleus of CN X) and sympathetics (intermediolateral cell column in the T1–L2 lateral horn of the spinal cord) and to the nucleus tractus solitarius. PVN parvocellular neurons also send axons to several important limbic-related structures, such as the amygdaloid nuclei, parabrachial nuclei, and locus coeruleus.
The PVN of the hypothalamus is a small region along the upper borders of the third ventricle in the dorsal hypothalamus. It contains a remarkable array of chemically specific neural populations. The magnocellular neurons produce oxytocin and vasopressin along with neurophysins and project to the median eminence. Some parvocellular neurons synthesize corticotropin-releasing hormone and send axons to the contact zone of the median eminence, where corticotropin- releasing hormone is released into the hypophyseal portal vessels. Parvocellular neurons also send descending projections to the brainstem (particularly the nucleus solitarius) and the intermediolateral cell column of the thoracolumbar spinal cord, where activation of the SNS can occur. The PVN therefore can coordinate the activation of both the neuroendocrine components (hypothalamic-pituitary-adrenal axis and cortisol secretion) and the autonomic components (sympathetic activation, diminished parasympathetic activity) of a stress response or activational response. The PVN receives inputs from many limbic regions and from brainstem sites (parabrachial nuclei, brainstem noradrenergic nuclei, nucleus tractus solitarius) that provide visceral information to the PVN. In addition, the PVN receives a variety of inputs that help it to monitor inflammatory mediators (IL-1β, IL-6, tumor necrosis factor [TNF]-α, prostaglandin E2 [PGE 2 ]), and other small molecules (nitric oxide) that reflect the outside chemical milieu. This information is received through the hypothalamus and circumventricular organs, and some of it through the vagus nerve afferents and nucleus tractus solitarius. Thus, PVN is a key regulatory site for behavioral responses that require autonomic reactivity.
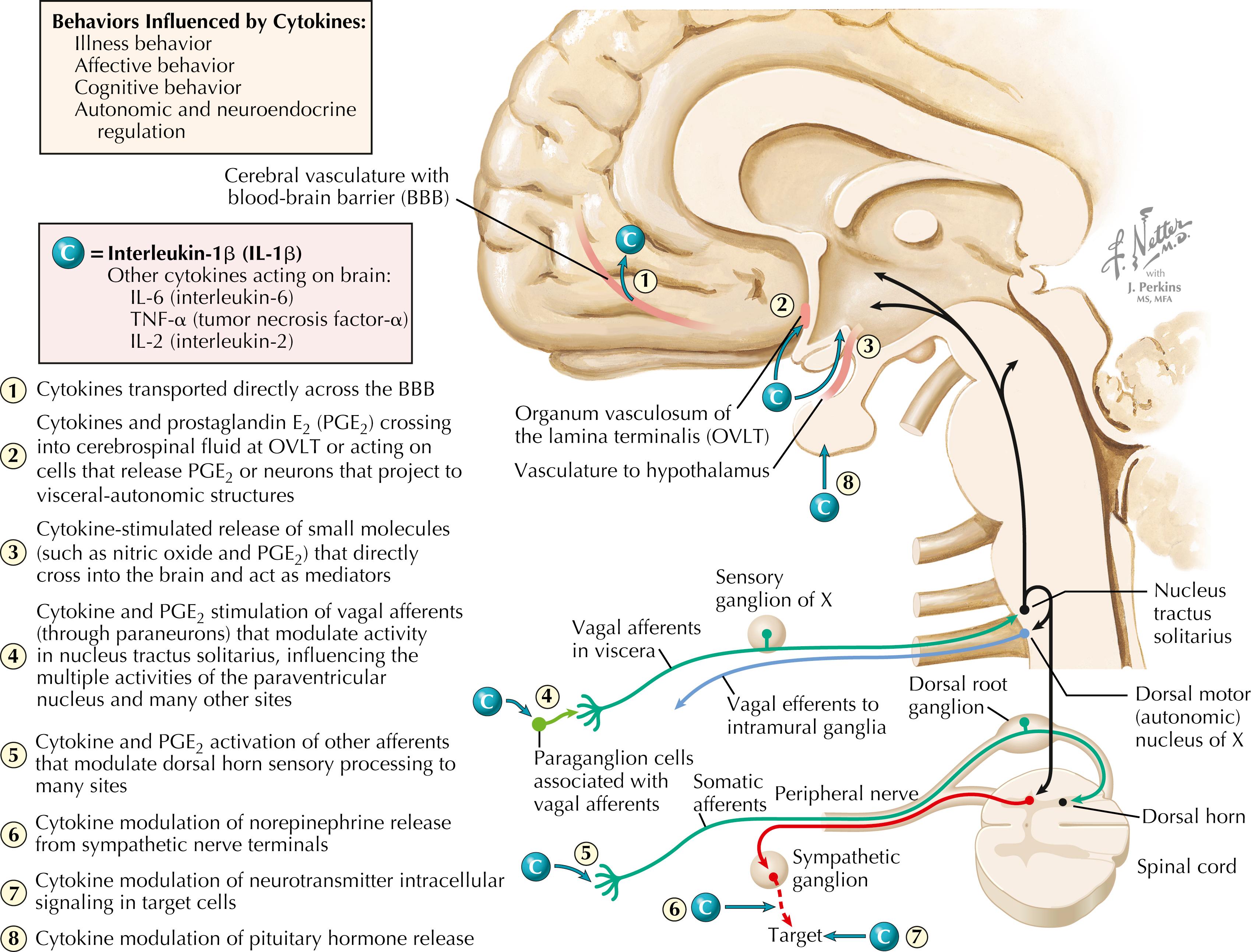
Cytokines, including IL-1β, IL-6, TNF-α, and IL-2, can influence central neuronal activity and behavior. This figure illustrates IL-1β access to the brain: (1) directly crossing the blood-brain barrier into the brain (especially in cortical regions); (2) acting on circumventricular organs (the OVLT) to release small mediators such as PGE 2 ; (3) acting on vascular endothelial cells to release nitric oxide, which acts in the CNS; (4) activating vagal afferents that project into the nucleus tractus solitarius via paraganglion cells; and (5) activating other afferent nerve fibers. IL-1β can evoke illness behavior (fever, induction of slow-wave sleep, decreased appetite, lethargy, classical illness symptoms), can influence autonomic and neuroendocrine regulation, and can influence both affective and cognitive functions and behavior.
There is a widespread influence of cytokines , especially inflammatory cytokines (IL-1β, IL-6, TNF-α), as well as prostaglandin E 2 (PGE 2 ), on the nervous system. A key target of these influences is the PVN of the hypothalamus. Inflammatory cytokines can provoke a robust activation of cortisol secretion (through the hypothalamo-pituitary-adrenal axis) and SNS activation (via descending projections of the PVN). The consequences of prolonged stress activation include increased risk for many chronic diseases, such as cardiovascular disease and stroke, metabolic syndrome, type II diabetes, and many cancers. The cytokines can influence the PVN and other central neurons through several mechanisms, including some direct transport into the forebrain, actions on neurons of the OVLT that release PGE 2 and signal the PVN, release of nitric oxide and PGE 2 from vascular endothelial cells, and activation of vagal afferents and other afferents that send neural signals to the PVN. Inflammatory cytokines also can stimulate the release of some hormones from pituitary cells, can alter neurotransmitter release in both the CNS and the autonomic nervous system (especially sympathetic norepinephrine), and can interact with neurotransmitter effects on target cells of autonomic innervation. Other cytokines such as IL-2 also appear to have central effects; the infusion of IL-2 in immunotherapy for some cancers was curtailed because of adverse effects of IL-2 on the brain, including depression and suicidal behavior.
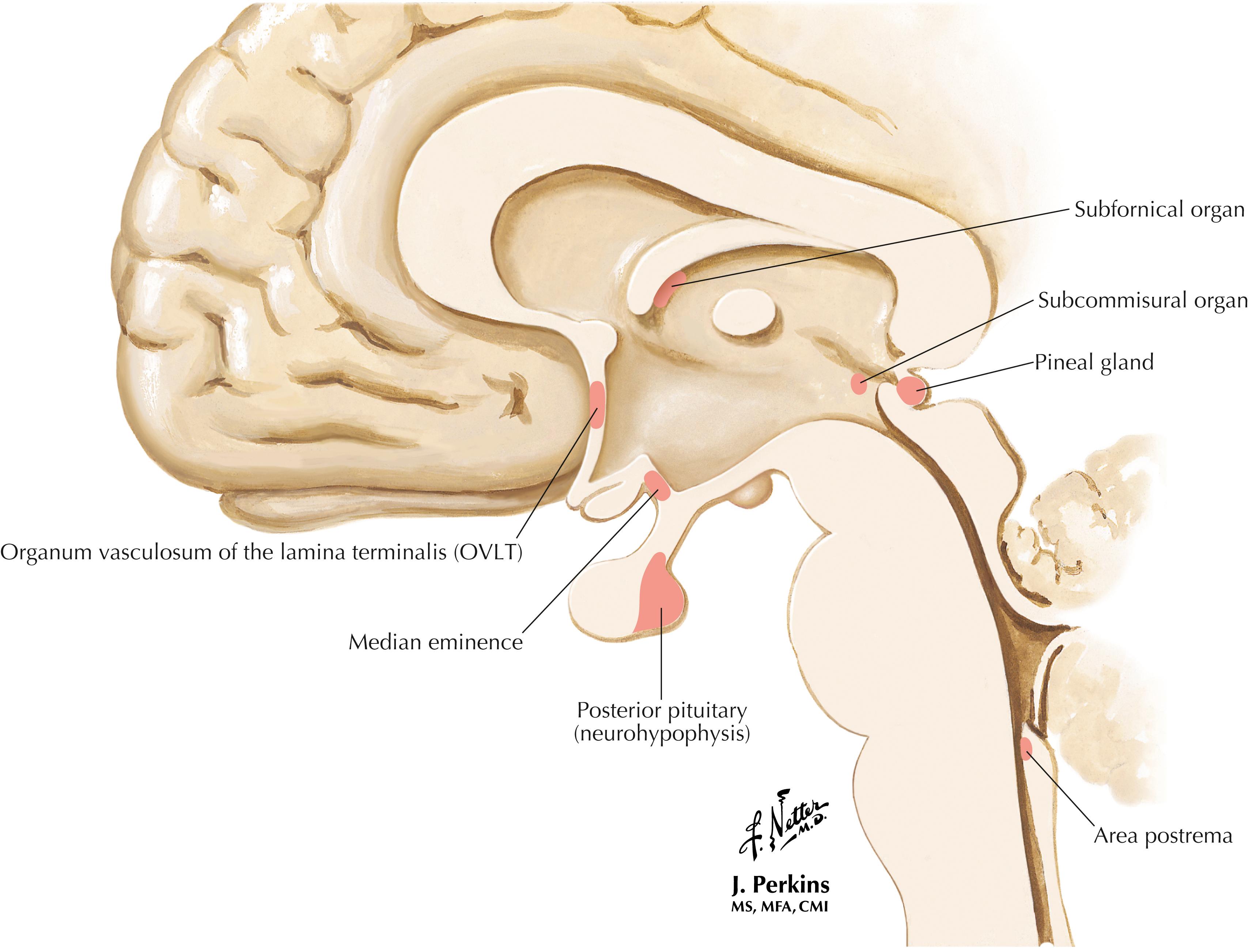
Circumventricular organs are “windows on the brain” that are devoid of the usual tight junction endothelial appositions and instead have fenestrated vasculature. Thus, the circumventricular organs have no blood-brain barrier. Some of these organs (the OVLT, the subfornical organ, and the area postrema) have associated neurons that project to the hypothalamic and other visceral structures. They also have cells that can release small molecules such as PGE 2 into the cerebrospinal fluid, thus affecting target structures at a distance. The neurohypophysis is a site of axonal release (from PVN and SON magnocellular neurons) of oxytocin and arginine vasopressin into the general circulation. The median eminence is a zone of neuroendocrine transduction for the secretion of releasing factors and inhibitory factors into the hypophyseal portal vasculature; these factors influence the release of anterior pituitary hormones. The pineal gland synthesizes and releases the hormone melatonin.
The CNS is protected from damage caused by many potentially harmful substances in the periphery by the blood-brain barrier. The CNS capillary endothelial cells contain tight junctions as well as specific transport mechanisms for the uptake of certain important substances (such as amino acids needed for neurotransmitter synthesis, glucose). Brain capillaries also can actively pump some substances out of the brain. Some regions of the brain contain fenestrated capillaries, and this permits the sampling of circulating substances. These are the circumventricular organs. The area postrema contains neurons that project to the nucleus tractus solitarius and activate the vomiting reflex. The subfornical organ contains neurons that respond to salt content in the blood and elicit protective neuroendocrine responses. The OVLT contains neurons that help to regulate blood pressure through an angiotensin II mechanism; these neurons also regulate PGE 2 availability to the PVN and other central areas to influence activation of the hypothalamo-pituitary-adrenal axis and the SNS. The OVLT and subfornical organ also respond to pyrogens and help to regulate hypothalamic responses for control of body temperature. At the median eminence, circulating hormones and other substances can interact with the projecting axonal terminals that secrete releasing hormones and inhibitory hormones at the contact zone for regulation of anterior pituitary hormonal secretion. The posterior pituitary and the pineal gland also have fenestrated capillaries, enabling their secretion of hormones directly into the systemic circulation.
Become a Clinical Tree membership for Full access and enjoy Unlimited articles
If you are a member. Log in here