Physical Address
304 North Cardinal St.
Dorchester Center, MA 02124

Why are we concerned with terminology? All research groups know what the words and terms they use mean.
It is possible that different research laboratories are finding the same transcription factors or extracellular matrix molecules and giving them different names. It is known that families of enzymes have in the past been renamed or renumbered when this has been identified. More recently, the importance of defining what is meant by commonly used terms has been realised, and many papers include their own definitions. The expansion of research and the names conferred on genes, proteins and cytokines means that it is prudent that everyone is explicit in what they include and attribute to the terms they use so that others can clearly see if they are also using these terms in the same manner.
Why include tables on flies and chick embryos?
Genomes of human and common laboratory animals have been published, and the proteins they encode are being investigated. The shared aspects of animal and human genomes provide information on evolutionary processes and, with environmental data, epigenetic processes as well. Common metabolic pathways can be explored in many species. Databases, research papers and the discussion on concepts derived from other species are therefore of value in the interpretation of human developmental processes.
Computers contribute greatly to developmental knowledge, but what are the concerns around the use of computers in developmental research?
It is true that the use of computer algorithms has permitted the analysis of gene products and led to the prediction of new proteins, which have been consequently verified. However, two concerns may still be considered: First, computers are now contributing directly to developmental databases without there always being input from a human curator to reflect on veracity. Second, many researchers within the computer side of development may have limited knowledge and understanding of developmental processes within an embryo and how the concepts of such knowledge have been derived. It is possible that we may not be able to revisit and advance the underlying theories of development. Yet at the same time, the complexity of research outcomes appears to be increasing.
Why does the appearance of a cell matter?
The genes, and therefore proteins, within a cell define its structure and function. The arrangements and interactions with other cells can alter the expression of genes within a cell. Two types of stable arrangements of cells into tissues are epithelia and mesenchyme. In an epithelium, the cells face the same direction and join together by junctions on the same types of cell borders. In mesenchyme, cells do not necessarily join at all but communicate through products they secrete into the space around them. The disposition of the intracellular organelles is different between epithelial cells and mesenchyme.
The plane of cell division within an epithelial sheet, controlled by polarity genes, can produce very different movements of the sheet itself. Interactions between epithelial cells and with mesenchymal cells are very specifically regulated.
Why are cell interactions important?
There is an agreed-upon terminology to explain types of interactions between cells and between tissues. An interaction termed instructive means that regardless of where it happens in a developing embryo, there is important information passing from one cell population to another. Some cell populations appear to project manage some local developmental domains.
Why is the space between embryonic cells important?
It used to be thought that the space between embryonic cells was just filled with water or some sort of hydrophilic molecules and fibres. This is still sometimes termed ‘ground substance’. This term is seriously misleading because the complexity of the extracellular matrix is now being realised and explored. The space between mesenchymal cells is filled by their secretions of collagen fibres, proteoglycans and very complex hydrophilic glycosaminoglycans. The cells also secrete a range of growth factors and cytokines into this matrix, which form gradients of stimulation to cells close to it or migrating through it, proving spatial and temporal developmental cues. The stiffness, degree of compression and mechanical movement of the matrix are also registered by mesenchyme cells, contributing to their interaction and responses.
Why are there two staging systems?
There are two staging systems because both the time at which an oocyte is released from an ovary, and when a particular blastocyst implants into the uterine endometrium is variable, the exact postfertilisation age of an embryo is difficult to establish. After the blastocyst has implanted, then factors it releases can be detected, and a pregnancy can be confirmed; however, the only clear date a woman may have for when she was not pregnant is the date of the start of her last menstrual period.
Obstetric timing of pregnancy is based on the time of the last menstrual period. Embryonic development is timed from fertilisation; this is about 14 days after the last menstrual period. Therefore the two systems always have a difference of 2 weeks. When an embryologist thinks of a 7-week embryo, it is very different from when an obstetrician thinks of a 7-week pregnancy. It is important that the extent of these differences is understood and the embryonic development in each case is known.
Why is knowing when organs and structures develop in an embryo important?
Rapidly dividing tissues are at risk if the mother takes medications or other drugs at these times. Many teratologic drugs or radiation therapies would be avoided if pregnancy was suspected. Furthermore, if medication is taken during pregnancy, its effects on organs can be predicted, and appropriate advice can be given to the patient.
Why is the age of the fetus so important?
During pregnancy, various biometric indices are measured to assess whether fetal growth is appropriate for gestational age or if it is affected by other factors such as placental insufficiency infection or diabetes. Fetuses that are small or large for gestational age may give cause for concern. It is important for biometric indices to be compared with relevant measurements. There is an ongoing debate whether this should be to those relevant for the country and culture of the mother or international set guidelines.
What criteria are used to determine causation in the field of teratology?
Methods for determining causation in the field of teratology use the Bradford Hill criteria. To consider the conclusion of causation, you do not need to satisfy all the criteria, but the more criteria that are satisfied, the more you can be confident that the association is causal.
What are the most common findings in congenital rubella syndrome?
The most common finding is deafness, which is associated with maternal infection during the first 16 weeks of pregnancy. Structural findings include congenital cataracts, patent ductus arteriosus and growth impairment. Cognitive impairment and encephalitis are also possible. It is estimated that 80% of fetuses are affected after maternal infection in the first trimester and 50% if the infection occurs between 12 and 16 weeks. After this gestation, adverse effects are rare; however, deafness has been reported.
What are the most common findings in isotretinoin-exposed fetuses?
Oral isotretinoin, a derivative of vitamin A, has been associated with a risk for spontaneous abortion, facial and palate defects, micrognathia, ear anomalies, conotruncal heart defects, thymic abnormalities as well as developmental delay. Importantly, cognitive effects can occur even in the absence of fetal brain abnormalities on ultrasound examination.
What are the criteria to diagnose a miscarriage in a first ultrasound scan?
An early embryonic demise should be suspected when the mean sac diameter is 25.0 mm or greater and there is no visible embryo or when the crown–rump length is 7.0 mm or greater and there is no visible cardiac activity. A second opinion on the viability of the pregnancy should always be sought before confirming the diagnosis to reduce the risk for misdiagnosis due to operator error.
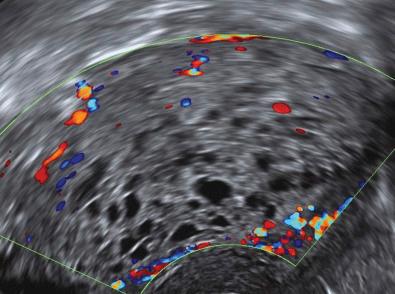
What is the diagnosis in the ultrasound scan image above?
It is a complete molar pregnancy that was confirmed on histology. Thick and cystic tissue within the uterine cavity without evidence of a gestation sac is highly suggestive of a complete molar pregnancy. Up to 95% of complete molar pregnancies are detected on ultrasound scan.
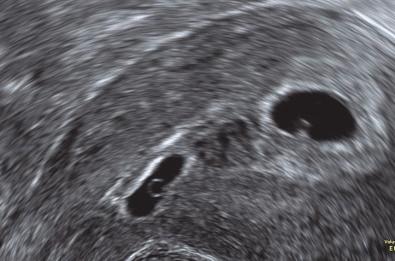
What is the diagnosis in the ultrasound scan image above?
It is a case of heterotopic pregnancy. A gestation sac is seen implanted normally within the uterine cavity (right). A second gestation sac is seen implanted anteriorly into a deficient caesarean section scar.
What is the evidence that the decidua is involved in controlling trophoblast invasion?
 are shown with anchoring cytotrophoblast cell columns and trophoblast invasion into the maternal decidua (bottom) . Maternal blood from decidual spiral arteries (A) fills the intervillous space in direct contact with syncytiotrophoblast (ST). Distinct trophoblast populations are shown. Villous trophoblast comprises: cytotrophoblast (CT) which forms the inner layer and is proliferative. CT fuse to form the outer syncytiotrophoblast layer (ST) which is in contact with maternal blood. Neither CT or ST stain for HLA-G. Extravillous trophoblast includes cytotrophoblast cell columns (COL), interstitial trophoblast (IT), endovascular trophoblast (ET) and placental bed giant cells (GCs); all stain strongly for HLA-G. Anchoring cell columns coalesce to form a continuous trophoblast shell (TS). From this shell, interstitial trophoblasts (ITs) invade through the decidual stroma to encircle and destroy the arterial media, which is replaced by fibrinoid material (F). ETs move in retrograde fashion down spiral arteries, displacing endothelial cells. On reaching the inner layer of the myometrium, trophoblast cells differentiate to multinuclear giant cells (GCs). The inset shows a representation of cellular interactions within the decidua. ITs are seen between large decidual stromal cells (S). Maternal leukocytes present are mainly uterine natural killer (uNK) cells with a few macrophages (Ms) and occasional T cells (Ts).")
In vivo and in vitro experiments show that extravillous trophoblast cells (EVTs) are inherently invasive. During normal placental development, EVTs migrate into the decidua as far as the myometrium ( eFig. 6.1 ). When the decidua is damaged, for example, if the placenta implants over a caesarean section scar, trophoblast invasion can proceed unchecked. This may result in conditions such as placenta accreta, in which EVT invades deep into the myometrium. Similar behaviour is seen in ectopic pregnancies when implantation occurs outside the uterine cavity in areas lacking decidua. This had led to the suggestion that the decidua plays an important role in regulating trophoblast invasion. Current evidence strongly suggests that uterine natural killer (uNK) cells in the decidua play a role in regulating trophoblast invasion; however, other cell types in decidua may also be involved. This role for natural killer (NK) cells is supported by data from a rodent model in which depletion of uNK cells during early pregnancy increased trophoblast invasion. [CR]
What mechanisms may protect the semi-allogeneic placenta from damage by maternal T cells?
In transplantation, allorecognition by T cells results in allograft rejection. The main drivers of this process are human leukocyte antigen (HLA)-A and -B molecules, neither of which are expressed by EVTs. There are two main sites of contact of the placenta with maternal T cells. Placental villi are in direct contact with maternal blood. The villi are lined by a continuous syncytiotrophoblast layer. This does not express major histocompatibility
 depends on their major histocompatibility (MHC) restriction and antigen specificity. A, CD8+ effector T cells may directly recognise paternal HLA-C, regardless of the antigen bound, if the paternal human leukocyte antigen (HLA)-C molecule structure is sufficiently different. B, CD8+ effector T cells may directly recognise maternal HLA-C on EVT presenting a paternal antigen. C, CD8+ T cells could indirectly recognise paternal antigen presented on maternal MHC class I on decidual antigen-presenting cells (dAPCs). This would rely on the process of cross-presentation. D, CD4+ effector T cells can indirectly recognise paternal antigen presented on maternal MHC class II on dAPCs. (Figure based on an idea by Emma Charlton.)")
(MHC) class I nor class II molecules so cannot be directly recognised by T-cell receptors on CD8 or CD4 T cells in maternal blood. A second site of contact occurs between EVTs that invade the decidua and encounter maternal T cells therein (see eFig. 6.2 ). EVT does not express MHC class II molecules, which eliminates direct recognition by maternal CD4+ T cells. EVTs do not have HLA-A or -B but do express HLA-E and HLA-G molecules which show minimal variation between individuals. This leaves HLA-C as the only polymorphic MHC class I expressed by EVTs which is capable of functioning as an alloantigen recognised by maternal T cells.
Pregnancies in which fetal and maternal HLA-C allotypes are mismatched show increased frequency of activated CD4+ T cells in the decidua. With no HLA class II expression on EVTs, CD4+ T cells can only recognise paternal antigens if presented on HLA class II molecules on maternal antigen presenting cells in the decidua. CD8+ T cells could theoretically recognise fetal antigens presented by HLA-C directly on EVT, but to date, there is no convincing evidence that CD8+ T cells specific for HLA-C are present in the decidua. Nor is it clear, whether if present, they result in any damage to placental cells. This has been directly tested in mice. No detrimental effects on pregnancy outcome occur in mouse models in which CD8+ and CD4+ T cells are activated against paternally inherited alloantigens known to be expressed on trophoblast. Several local mechanisms appear to contribute to avoiding damaging responses to trophoblast by promoting a tolerogenic environment in the decidua. These include silencing of chemokine genes in decidua; this prevents T cells entering decidual tissue and the expression of tolerogenic cytokines such as interleukin-10 and transforming growth factor-beta. In summary, there is no compelling evidence to date that abnormal T-cell responses are associated with human pregnancy failure. A full account of this complex topic is available in the excellent review by Nancy and Erlebacher. [CR]
Can measurement of phenotype and function of immune cells in blood give useful information about whether immune cells in the decidua may be able to target the placenta?
During pregnancy, cytotoxic T cells are found in blood with specificity for fetal minor histocompatibility antigens as well as inherited paternal HLA. These responses are generated due to fetal cells that have escaped from the placenta into the maternal circulation. However, the lack of expression of HLA class I or class II by syncytiotrophoblast means that fetal antigen-specific T cells circulating in the blood cannot target placental syncytium directly. Even if some fetal-specific T cells do reenter the decidua, there is no clear evidence they can target EVTs because most have been shown to be restricted to recognition of HLA-A or -B, neither of which are expressed by EVTs. [CR]
Many studies have also shown considerable phenotypic and functional differences in both T cell and NK subtypes between cells isolated from peripheral blood and those in the decidua caused by the local microenvironment. Finally, there is evidence that many NK cells and macrophages in the decidua may be tissue resident and not even derived from precursors circulating in peripheral blood. For these reasons, it seems highly unlikely that studies of immune cells in blood can give useful insights into the extent of maternofetal tolerance in the decidua. A more detailed discussion of this topic is available in Moffett and Colucci. [CR]
Chakraborty D., Rumi M.A., Konno T., Soares M.J. Natural killer cells direct hemochorial placentation by regulating hypoxia-inducible factor dependent trophoblast lineage decisions. Proc Natl Acad Sci U S A . 2011;108(39):16295–16300.
Nancy P., Erlebacher A. T cell behavior at the maternal-fetal interface. Int J Dev Biol . 2014;58:189–198.
Verdijk R.M.1, Kloosterman A., Pool J., et al. Pregnancy induces minor histocompatibility antigen-specific cytotoxic T cells: implications for stem cell transplantation and immunotherapy. Blood. 2004;103(5):1961–1964.
Moffett A., Colucci F. Uterine NK cells: active regulators at the maternal-fetal interface. J Clin Invest. 2014;124:1872–1879.
What are the key features of the normal term placenta from the fetal side?
:1, 2012.")
The placenta is a disclike circular organ usually inspected in its ‘inverted’ state from the fetal, or chorionic plate, surface.
The umbilical arteries branch out on the chorionic plate, each penetrating the plate and enter the stem (truncus) of a villous tree; each is accompanied by a chorionic plate vein draining oxygenated and nutrient-rich blood back towards the umbilical vein.
The amniotic membrane covers the chorionic plate. At the placental margins, the trophoblast and associated mesenchyme of the chorionic plate fuses with the basal plate to become the chorion laeve, often described as the fetal membranes ( eFig. 7.1 ).
What are the key features of the normal term placenta from the maternal side?
The basal plate of the delivered placenta is a heterogeneous mixture of trophoblastic and decidual cells, embedded in large amounts of extracellular debris, fibrinoid and blood clot.
The basal surface is divided into ‘maternal lobes’ of the placenta. Within these are the functional units of the placenta, termed ‘placentomes’, each of which consists of three or four placental villous trees perfused centrally by a spiral artery.
The mature human placenta comprises 15 to 20 placental lobules (two or more primary stem villi and corresponding secondary and tertiary branches) and thus up to 50 placentomes, suspended in the intervillous space and perfused by maternal blood at high-flow and low pressure.
Describe the macroscopic abnormalities of the placenta.


These occur in 10% of term cases and are identified from the fetal side of the placenta.
These include succenturiate lobes, velamentous cord insertion or circumvallate margins, where the ring is undergrown by villous trees and the intervillous space and the membranes insert medial to the edge of the chorionic plate ( eFig. 7.2 ).
Second trimester ultrasound can identify some gross abnormalities of the placenta; however, these are not indicative of placental efficiency, especially if other aspects of placentation, such as uteroplacental blood flow are normal and there are no maternal comorbidities.
At delivery, the fetal surface of the placenta may show evidence of intraamniotic infection, such as cloudy yellow malodorous discolouration or chronic staining by meconium when clinically relevant ( eFig. 7.3 ).
How is the maternal uteroplacental circulation of the intervillous space established by the human haemochorial placenta?
The uteroplacental circulation is prepared at the outset of pregnancy by the coordinated interactions of the implanting placenta and the maternal endometrium.
The endometrial spiral arteries in the uterine wall grow and become convoluted under the control of pre- and postovulation hormones.
Before menstruation, these vessels constrict causing foci of endometrial ischemia, necrosis and haemorrhage.
However, when pregnancy occurs, the persisting progesterone expression by the corpus luteum maintains the vascular development, initiating decidualisation.
Decidualisation involves the endometrial fibroblast differentiating into large secretory stromal cells, with an influx of specialised pro–tissue remodelling maternal immune cells.
During blastocyst attachment and invasion of the decidua, the trophectoderm proliferates rapidly to form the trophoblastic shell that lines the developing intervillous space.
Next the primary chorionic villi of the placenta form via fusion of these trophoblast shell cells into syncytiotrophoblast and lacunae formation.
Most villi commit to the villous placenta while the rest form the anchoring chorionic villi that attach the placenta to the uterus. The anchoring cytotrophoblast columns at the outermost edge of the trophoblast shell give rise to the highly invasive mononuclear extravillous trophoblasts (EVTs), which do not proliferate after they have exited the anchoring column.
The EVTs penetrate the maternal decidua and encounter the small uterine capillaries, which are then blocked by the formation of an endovascular plug of EVT, thereby preventing maternal blood flow and oxidative damage to the embryo and placenta in the first few weeks of development.
During this time, the developing embryo is fed by histiotrophic nutrition by endometrial glandular secretions.
Upstream of the EVT plug, there is a progressive transformation of the maternal spiral arteries involving the loss of the vascular smooth muscle and endothelium. This process is started by decidual natural killer and macrophage cells clustering around and in vascular walls of the spiral artery, secreting angiogenic growth factors and matrix metalloproteases, which break down the extracellular matrix of the vessel wall and contribute to the loss of the vascular smooth muscle cells and consequent widening of the vessel lumen. (Please see Fig. 7.4 in Chapter 7.)
Between 8 and 12 weeks of gestation, loss of EVTs from the endovascular plug destabilises it and initiates maternal blood flow into the intervillous space.
By 16 to 18 weeks of gestation, interstitial EVTs in the decidua and endovascular EVTs migrate along the vessel lumen as far as the first third of the myometrium.
The EVTs gradually reline the transformed spiral arterioles, acquiring some endothelial characteristics as they do so and restabilising the enlarged tube by depositing fibroid.
These factors turn the uterine spiral arteries into flaccid tubes that are unable to contract because of the absence of smooth muscle and result in a high flow low-pressure system that is characteristic of the uteroplacental system.
Defects in the transformation of the uterine spiral arteries have been implicated in a range of placental complications of pregnancy, from chromosomally normal late first trimester miscarriage to preterm birth, intrauterine growth restriction and preeclampsia.
How does the differentiation of the placental villous tree maximise the transport of oxygen, nutrients and waste between the maternal and fetal circulations?
Differentiated villous trophoblast buds, invaded by mesenchymal cells, undergo vasculogenesis (the de novo formation of blood vessels) and are termed primitive ‘mesenchymal villi’. They are the first tertiary villi (containing three layers – trophoblast, fetal mesenchymal cells and developing blood vessels).
New mesenchymal villi are formed by vascularisation of trophoblastic sprouts from existing villi, thereby expanding the villous trees. During the second trimester, the more mature villi become the stem villi, characterised by mature arterioles and venules within a fibrosed stromal core.
The peripheral capillaries of the villous stroma are retained as the paravascular capillary network, but by term, the larger stem villi possess very few paravascular capillaries because they are nonfunctional.
In the third trimester, the villous growth changes to form the terminal villi, the functional unit of the human placenta.
From 26 weeks onwards, at the tips of the vascular tree, the immature intermediate villi differentiate to form differentiated structures, the mature intermediate villi (MIV), which are the source of the functional gas-exchanging terminal villi (TV).
TV form from the rapid capillary elongation and lateral prolapse at multiple sites.
This outgrowth of TV capillaries occurs at a rate much greater than the rate of trophoblast proliferation and fusion, leading to a very thin and anuclear ‘vasculo-syncytial membrane’ (VSM) that maximises the diffusion-limited transfer of oxygen.
The third trimester of pregnancy is characterised by an exponential rise in the formation of TV and accompanying VSM, resulting in a large surface area (14 m 2 ) to accommodate the diffusional needs of the growing fetus.
Together with increasing fetal blood pressure, these anatomical changes in the structure of the placental villi result in around 40% of fetal cardiac output entering the umbilical arteries.
The terminal villi are visible as grape-like outgrowths from the surface of the mature intermediate villi. ( eFig. 7.4A ) shows the fetal capillaries coiling inside the terminal villi, stained for endothelial cell marker platelet endothelial cell adhesion molecule-1 [PECAM-1].) Electron microscopy of terminal villi ( Fig. e7.4B ) demonstrates these features, with sinusoidal dilated capillaries that protrude to the surface of the syncytiotrophoblast forming the VSM.
List the exchange mechanisms and pathways available by which solutes may be transferred across the placenta.
These are diffusion and filtration (pores), diffusion (pores or channels), facilitated diffusion (carriers), secondary active transport (carriers including co-transporters and exchangers), primary active transport (pumps) and endocytosis and exocytosis (both vesicles).

What happens to maternofetal O 2 transfer after a drop in umbilical artery pO 2 , and what is the mechanism underpinning this?
Fetal blood has a higher haemoglobin concentration than maternal blood, and maternal haemoglobin has a lower affinity for O 2 than fetal haemoglobin, so the single most important determinant of O 2 transfer from maternal to fetal blood is the O 2 content of blood perfusing the placenta in the umbilical arteries. Presuming uterine artery pO 2 remains constant, maternofetal O 2 transfer will increase because of the greater diffusion gradient. Compensatory changes in fractional O 2 extraction keep fetal O 2 uptake relatively constant during short-term changes in umbilical and uterine blood flow
Therapeutics, proposed to increase blood flow to the placenta, have been suggested as potential treatments for fetal growth restriction (FGR). Are these types of therapeutics likely to directly increase the placental transfer of amino acids?
No, amino acids are hydrophilic and have a low permeability across the placenta. Thus changes in blood flow are unlikely to affect transfer of amino acids significantly, whose transfer is said to be ‘membrane’ or ‘diffusion’ limited.
In FGR, what happens to the expression of the mechanistic target of rapamycin (mTOR) in a placenta, and why is it potentially important for placental transfer of amino acids?
Placental mTOR acts as a nutrient sensor and has reduced expression in FGR. Studies on placental tissue and cytotrophoblast cells in vitro show that inhibiting mTOR activity results in reduced amino acid transporter activity. Thus reduced expression of mTOR in FGR may have significant consequences for the delivery of amino acids to the fetus.
Describe the major mechanisms by which Ca 2+ is transferred, transcellularly, across the placenta from maternal to fetal blood. Name the proteins involved and their roles.
Ca 2+ enters the syncytiotrophoblast, across the maternal-facing microvillous plasma membrane (MVM), down an electrochemical gradient from maternal blood. This diffusion is likely to involve channels located in the MVM, with transient potential vanilloid type 6 (TRPV6) one protein implicated. Once inside the cytosol of the trophoblast, Ca 2+ binds to a calcium binding protein, with calbindin-D 9K . Ca 2+ efflux occurs from the cytosol of the trophoblast, across the basal membrane (BM) (and hence fetal capillary endothelium) and into fetal blood. This process occurs against an electrochemical gradient and so requires adenosine triphosphate (ATP) in the form of the plasma membrane calcium ATPase (plasma membrane Ca2+ ATPase [PMCA] or Ca 2+ ATPase or calcium pump), located on the BM. PMCA in particular ensures that fetal blood is hypercalcaemic compared with maternal blood.
What are the sonographic diagnostic features of Breus mole?
Sonographic features include noncompressible echogenic layers, at least 1cm deep and occupying at least 50% of the chorionic plate surface (due to laminated maternal–fetal haemorrhage) between the chorionic plate and the placental parenchyma. Colour Doppler detects no flow within the mass, and the surface chorionic plate vessels and umbilical cord are easily identified on the fetal surface adjacent to the amniotic fluid.
Is fetal survival possible with this diagnosis?
The fetal survival rate was 50% in a recent series and was predicted by normal umbilical artery Doppler. [CR]
How does a chorioangioma affect a developing fetus?
A large chorioangioma may induce cardiac failure, presenting as reduced fetal movements with nonimmune hydrops, which in the worse cases can lead to fetal death The placenta may show villous oedema and there may be ‘mirror syndrome’ type of preeclampsia.
Can fetal prognosis be improved by any in utero intervention?
Answer 4
Yes. Rarely, depending on gestational age, access and severity of fetal compromise, a large chorioangioma may be embolised, or its feeding fetal vessels may be ablated by fetoscopic laser. 2
What are the common risk factors for invasive placentation accompanying placenta previa?
Any type of uterine scarring that may result in deficient areas of endometrium and myometrium can cause invasive placentation. The most common cause is previous caesarean section; others include open myomectomy, multiple dilation and curettage procedures and hysteroscopic surgery with Asherman syndrome.
What are the key diagnostic features of placenta previa increta on ultrasonography or magnetic resonance imaging (MRI)?
These features are loss of normal lower segment myometrium (dehiscence) replaced by an expanded area of vascular lacunae, projection of the base of the placenta into the posterior wall of the full bladder and multiple lacunae within the placenta (dark intraplacental bands on MRI) that are fed by aberrant maternal arterioles. 3
What are the causes of an abnormally thick placenta (>4 cm) on ultrasound examination after 20 week’s gestation?
Any cause of fetal anaemia (immune and nonimmune, fetomaternal haemorrhage)
Any cause of fetal cardiac failure (e.g. fetal cardiac arrhythmia)
Placental tumour (vascular chorioangioma)
Massive thrombohaematoma (Breus mole)
Central retroplacental haematoma (abruption)
Maternal anaemia or high-altitude pregnancy
Subamniotic haemorrhage
Placental hyperinflation
What are the features and pregnancy risks with placental hyperinflation?
The features of placental hyperinflation are a second trimester finding of a small and thick (thickness >50% of length) placenta, typically associated with a combination of low maternal pregnancy-associated placental protein A (PAPP-A) and high α-fetoprotein levels. Early-onset intrauterine growth restriction (IUGR) may evolve, with abnormal umbilical artery Doppler. Therefore the risks are a need for early delivery by caesarean section and that if unrecognised, that stillbirth may occur because of undiagnosed IUGR.
Why is this different from Breus mole?
In placental hyperinflation, the widespread distal villous hypoplasia is likely associated with reduced density of the normal anchoring villi, which normally maintain parallel alignment of the chorionic and basal plates of the placenta in the face of maternal perfusion pressure from the spiral arterioles. Thus when the placental villi are not adequately anchored to the basal plate, the placental intervillous space expands. Note that the excess maternal blood is still within the intervillous space, which is why the placenta in utero is wobbly or compressible by the ultrasound probe. By contrast, in Breus mole, blood is abnormally sited (haemorrhage) outside of the normal vascular space and is thus noncompressible. After delivery, a Breus mole placenta remains thick because of organisation of its haematoma.
Note that in both diseases, umbilical artery Doppler and gestational age at presentation are predictors of fetal survival.
What are the associated risk factors for vasa previa?
The risk factors for vasa praevia include a succenturiate lobe, minor placenta previa or a velamentous cord insertion.
What is the role of transvaginal colour Doppler?
This modality of imaging gives high resolution of structures within 3 to 4 cm of the internal os and assesses cervical length accurately. In grey-scale real-time imaging, fetally derived vessels may be seen running on the amniotic membrane surface. Using low-flow (10–15 cm/sec) colour velocity settings, the fetal umbilical arteries or vein can be demonstrated.
What is the management of vasa previa?
Planned caesarean delivery (e.g. at 37 weeks’ gestation) avoids the risk for rupture of these vessels that likely would occur during normal labour, especially with amniotomy. Women with a history of preterm birth or a short cervix may be admitted to hospital until planned caesarean delivery, such that the operation can be done immediately if labour occurs or the membranes rupture spontaneously. 4
Alanjari A., Wright E., Keating S., et al. Prenatal diagnosis, clinical outcomes, and associated pathology in pregnancies complicated by massive subchorionic thrombohematoma (Breus’ mole). Prenat Diagn . 2013;33(10):973–978.
Hosseinzadeh P., Shamshirsaz A.A., Javadian P., et al. Prenatal therapy of large placental chorioangiomas: case report and review of the literature. AJP Rep . 2015;5(2):e196–e202.
Jauniaux E., Collins S.L., Jurkovic D., Burton G. Accreta placentation: a systematic review of prenatal ultrasound imaging and grading of villous invasiveness. Am J Obstet Gynecol . 2016;215(6):712–721.
Swank M.L., Garite T.J., Maurel K., et al. Vasa previa: diagnosis and management. Am J Obstet Gynecol . 2016;215(2):223.e1–e6.
Describe how the interatrial septum forms and how this explains the morphology of secundum atrial septal defect.
 grows between the right and left atria. Its lower border grows towards the atrioventricular cushion, the gap between them forming the ostium primum ( B ).")
The normal interatrial septum has several components. Much of what appears to be septum is actually infolding of the extracardiac tissue between the two atria. The true septum consists of the oval fossa and its rim. The fossa is closed by a membrane, the flap valve, that is not completely sealed in 25% of the population. The flap valve is attached on the left side and viewed from the left atrium the oval fossa is not visible as an oval depression. The remainder of the party wall of the two atria is formed by atrial muscle and adipose tissue between the lower border of the oval fossa and the atrioventricular junction.
The atria develop as paired outgrowths from the caudal part of the heart tube cephalad to the sinus venous. The space between the atria sees the growth of a primary septum (the septum primum) that fuses with the atrioventricular cushions. There is a mesenchymal cap on the edge of the primary septum that is continuous ventrally with the ventral atrioventricular cushion and dorsally with the dorsal mesenchymal protrusion and the dorsal atrioventricular cushion. The gap between the free edge of the growing primary septum and the atrioventricular cushions is called the ostium primum ( eFig. 10.1A and B ). Before fusion of the septum primum and the atrioventricular cushions is complete (at about day 38), the septum breaks down in its ventral part to form a foramen secundum, thus continuing to permit blood to pass between the two atria ( eFig. 10.1C ). A second septum – the septum secundum – grows on the right side of the septum primum beginning at about day 41, by infolding of the muscular wall of the atrium, and partly closes the defect ( eFig. 10.1D ). The septum secundum is thicker and more muscular than the septum primum. It grows downwards, and its anterior part fuses with the endocardial cushions, but a defect remains – the oval foramen.
The right side of the atrial mass incorporates the right horn of the sinus venosus and forms a pair of valves around its orifice. Fusion of the anterior part of these valves creates the septum spurium, which contributes to closure of the atria. The left valve of the sinus venosus is incorporated into the developing septum secundum, but occasionally, remnants of it persist as thin threads attached to the right side of the septum.
Throughout fetal life, blood can pass from the right to the left atrium via the oval fossa and ostium secundum. After birth when the left atrial pressure rises because of increased pulmonary venous return to the left atrium, the flap valve is pushed against the septum and in more than 75% of cases seals by fibrosis. In 25% of individuals the flap remains unsealed and potentially openable – the so-called persistent foramen ovale. An atrial septal defect of secundum type occurs when the flap valve is insufficient in size to close the oval fossa under normal circumstances. The defect is usually small and situated anterosuperiorly ( eFig. 10.1E ), but multiple defects may be present ( eFig. 10.1f ), the defect may be large and the entire flap valve may be absent. Any interatrial communication outside the oval fossa is by definition not a secundum atrial septal defect. Such defects occur around the coronary sinus, the atrioventricular septum or adjacent to the superior caval vein.
Describe how the development of the pericardium results in the normal pericardial anatomy.
The pericardial cavity is a serous-lined cavity surrounding the heart. It has an outer fibrous layer and an inner serous layer. The fibrous layer is attached to the great vessels that anchor the heart in the chest. The serous layer covers the epicardial surface of the heart and the inner aspect of the fibrous pericardium, being reflected onto both at the attachments. There are several unique anatomical features of the pericardium that are explained by the embryology of the heart.
The cavity of the pericardium is a result of the development of the cavity of the embryo. The folding of the embryo results in the heart tube being projected into that cavity with a dorsal attachment to the body wall – the dorsal mesocardium. The epicardial cells derive from the polyp-like proepicardium near the transverse sinus. The dorsal mesocardium breaks down, leaving only the attachments of the two vascular poles, thus forming the transverse pericardial sinus. When the heart tube undergoes looping, there is a loop to the right and a fold anteriorly. This results in the atria being positioned posteriorly and the arterial outflow anteriorly and the transverse sinus of the pericardial cavity lying between these two structures. Folding also results in a recess of the pericardial cavity behind the atria. Because of the incorporation of the pulmonary venous return into the developing atrium, the original single vein is incorporated, together with its branches, which extends the pericardial attachment to the left, thus forming the blind ending space known as the oblique sinus ( eFig. 10.2A and B ).
The original left cardinal vein regresses, but in many cases, a fold of pericardium persists at its site extending to the site of the oblique vein of the left atrium ( eFig. 10.2C ). If there is a persistent left superior caval vein, it follows this course.
. The interatrial junction stretches horizontally to the left of the picture. The pulmonary veins are evident at the corners of the left atrium. Extending from the inferior caval vein to the pulmonary veins, the cut surface of the pericardial attachment can be seen. These attachments delimit the oblique sinus of the pericardium.")
Describe how the embryology of the thoracic arterial system helps understand how arterial vascular rings develop.
There is a complex process or modelling of the arteries of the head, neck and thorax. The outflow of the heart has a pair of dorsal aortae that fuse below the level of the fourth thoracic segment and become connected with the umbilical arteries. The pharyngeal arches, as they develop, acquire an arterial supply bilaterally—the aortic arches. The first pair of arches develops from folding of the embryo during the fourth week (days 22–24), causing the paired dorsal aortae attached to the cranial end of the heart to form a pair of dorsoventral loops. Between days 26 and 29, four additional pairs of aortic arches develop in succession from cranial to caudal within the mesenchyme of the pharyngeal arches, connecting the aortic sac at the superior end of the truncus arteriosus to the dorsal aortae; arches 2, 3, 4 and 6. The fifth aortic arch never develops. The first two pairs of arch arteries regress, and the later arch arteries are forming. Aortic arch arteries 3, 4 and 6 thus give rise to the arteries of the head, neck and thorax ( eFig. 10.3A and B ). The paired third arch arteries persist as the proximal parts of the internal carotid arteries (the distal parts being the cephalad extension of the dorsal aortae). The fourth and sixth arches remodel asymmetrically with more of the left-sided structures persisting than on the right. The fourth arches give rise to the origins of the subclavian arteries, and the sixth arches give rise to the proximal pulmonary arteries. On the left side the fourth arch gives rise to the arch of the aorta (hence the normal left-sided aortic arch!). The greater parts of the subclavian arteries actually derive from the right and left seventh intersegmental arteries which connect with the remnants of the fourth arches. The sixth arch on the left side gives rise to the arterial duct (a normally left-sided structure!) ( eFig. 10.4C and D ).
In the normal situation, the parts of the dorsal aortae connecting the third and fourth arches on both sides disappear. On the right side, the part of the right dorsal aorta connecting the fourth and sixth arches also disappears as well as the connection between the sixth arch and the fused dorsal aortae. On the left side, this part of the dorsal aorta persists as the descending aorta. The aortic sac gives rise to the proximal aortic arch and brachiocephalic artery. Failure of regression of any of these segments can result in a vascular ring around the oesophagus and trachea.
 and the dorsal aspect ( B ). The folded heart tube is seen in the centre of the picture. From the aortic sac, three pairs of aortic arches arise, from superior to inferior: arch 3, arch 4 and arch 6. Arches 1 and 2 have already involuted. The arches connect to the paired dorsal aortae that are fused in their caudal parts. It is from arches 3, 4 and 6 with their associated parts of dorsal aortae that the definitive arterial system of the thorax develops. The three pairs of veins entering the horns of the sinus venosus are represented in blue. From lateral to medial they are the cardinal veins, umbilical veins and vitelline veins.")
Explain in terms of their embryologic development why a persistent left superior caval vein drains to the coronary sinus.
Despite opening into the right atrium, the coronary sinus is a left-sided structure. This is supported by the fact that in right atrial isomerism, the coronary sinus is absent.
The coronary sinus derives from the left horn of the sinus venosus. In the developing venous system, there are three pairs of major veins that drain to the respective horns of the sinus venosus. They are from medial to lateral: right and left vitelline veins (from the yolk sac), right and left umbilical veins (from the placenta) and right and left common cardinal veins. The common cardinal veins are formed by confluence of the anterior and posterior cardinal veins, the anterior cardinal veins draining the head region.
With growth of the atria, the right horn of the sinus venosus is incorporated into the right atrium to form the sinus venarum (the smooth-walled component of the right atrium).
During the fourth and fifth weeks of development, the venous system undergoes a left-to-right shunting of blood. The left vitelline vein involutes with consequent shrinkage of the left horn of the sinus venosus. By 10 weeks’ gestation, the left common cardinal vein is obliterated, leaving the coronary sinus and the oblique vein of the left atrium as remnants.
If the connection fails to regress, then a persistent left superior caval vein drains to the coronary sinus. An innominate vein may or may not be present. Because of the increased blood flow, the coronary sinus is larger than usual.
 and dorsal ( B ) aspects and at 35 days’ development ( C ) (viewed from behind). The venous pole of the heart is inferior and shows right and left horns of the sinus venosus that are symmetrical and receive the three major veins (from medial to lateral: vitelline vein, umbilical vein and common cardinal vein). The common cardinal veins are formed by the confluence of the anterior and posterior cardinal veins, the former draining the head region. At this age, the junction of the sinus venosus with the common atrium is broad and central. By 35 days’ gestation, there has been major remodelling of the venous system. The left umbilical and vitelline veins have lost or will very soon lose their connection to the sinus, with only the common cardinal vein draining to the left horn. The left horn will become the coronary sinus, and by 10 weeks’ gestation, the left common cardinal vein will become obliterated with only a small remnant – the oblique vein of the left atrium. The right horn of the coronary sinus has shifted to the right side and is incorporated into the wall of the right atrium. The superior caval vein is formed from the right common cardinal vein. The hepatorenal part of the inferior caval vein develops from the right vitelline vein.")
What is the clinical and experimental evidence that lung expansion is a critical stimulus for fetal lung growth?
Conditions that either prevent the fetal lungs from expanding, such as congenital diaphragmatic hernias or pleural fluid accumulations (fetal hydrothorax), or cause compression of the fetal chest, such as oligohydramnios, or impair fetal skeletal muscle activity or bone development all result in fetal lung hypoplasia because of the inability of the lungs to fully expand. Conversely, clinical conditions such as laryngeal or tracheal atresia prevent the normal flow of liquid out of the lungs result in increased fetal lung expansion. These lungs are hyperplastic with increased alveolarisation.
Experimentally, this has been demonstrated in sheep, in which alterations in the volume of lung liquid result in profound changes in lung growth and maturation. Prolonged lung liquid drainage causes severe hypoplasia and a cessation of lung growth. In contrast, experimentally occluding the trachea, so that the volume of liquid within the lungs increases, results in increased lung growth and maturation.
What is the evidence that the transpulmonary hydrostatic pressure drives lung liquid clearance after birth rather than activation of amiloride-blockable sodium channels?
There has been considerable debate in the literature about the mechanisms behind airway liquid clearance at birth and the timing at which this process commences. 1-3 Nevertheless, when taken altogether (see later), it is evident that airway liquid clearance can occur due to a variety of different mechanisms. [CR] However, in any one infant, the mechanism that provides the greatest contribution will likely differ depending on the timing (gestational age) and mode of delivery (vaginal vs caesarean section delivery).
Although it has been suggested that airway liquid volumes can decrease days before birth, 4,5 this has not been confirmed in pregnancies with established normal amniotic fluid volumes. 6 Oligohydramnios is known to reduce lung liquid volumes because of an increase in transpulmonary pressure, causing the loss of lung liquid. 7 Similarly, any situation that reduces the available intrauterine space, such as the presence of a twin, may reduce lung liquid volumes before labour onset. However, when considered in relation to the lung’s capacity to clear airway liquid after birth (see later), this debate appears somewhat esoteric unless the infant is delivered by caesarean section. In this situation, the mechanisms for airway liquid clearance during birth are absent, necessitating that all airway liquid is cleared across the airway epithelium, with little being lost via the nose and mouth. 8
Fetal postural changes: Numerous publications have described the loss of large volumes of liquid via the nose and mouth after delivery of the infant’s head. 1,9 This mechanism of lung liquid loss is commonly referred to as ‘vaginal squeeze’. However, this description is not entirely accurate because the infant’s chest offers little resistance to delivery compared with the head and shoulders. 10 Instead, it is thought that uterine contractions force a change in fetal posture, which greatly increases fetal spinal flexion, 7 while oligohydramnios increases abdominal pressure, elevating the diaphragm and increasing transpulmonary pressure, resulting in lung liquid loss via the nose and mouth. This process likely explains the ‘gushes’ of liquid that have been observed after delivery of the infant’s head.
Na + reabsorption: Until recently, the primary mechanism responsible for airway liquid clearance at birth was thought to result from Na + uptake across the airway epithelium, which reverses the osmotic gradient leading to airway liquid reabsorption. 4 This mechanism is stimulated by increased circulating adrenaline and vasopressin levels, released in response to the stress of labour, and provides a convenient explanation for why infants born by caesarean section have a higher risk for transient tachypnea of the newborn (wet lung). 4,8 However, this mechanism only develops in late gestation and is absent in the immature lung of preterm infants because RNA transcripts encoding epithelial Na channel (ENaC) subunits are virtually undetectable in the immature human lung. 11 Clearly, preterm infants cannot use this mechanism to clear airway liquid. 12,13
Although adrenaline and vasopressin infusions inhibit lung liquid secretion and activate liquid reabsorption late in gestation, 14-16 pharmacological doses are required to achieve liquid reabsorption. 15,17 Furthermore, based on the measured reabsorption rates achieved, it would take hours to clear all liquid , 14-16 and adrenaline would have to remain elevated throughout this time. 18 However, normal healthy infants can clear their airways of liquid and establish effective gas exchange within seconds to minutes of birth. 13,19,20 Similarly, although heart rates commonly increase (from ∼100 to 160 beats/min) after birth, 21 there is no evidence of a sustained tachycardia, 21 which would be expected during a sustained stress-induced catecholamine release.
Although there is considerable evidence indicating that ENaCs help to control airway liquid in an aerated lung, 11,22 their role in airway liquid clearance at birth is less certain. Indeed, although inhibiting ENaC activity with amiloride delays lung liquid clearance in spontaneously breathing newborns, it does not prevent it and its effect in ventilated newborns is restricted to the expiratory phase. 23 ENaCs are encoded by three different genes (α-, β- and γ-), with the mature protein composed of three subunits. 18 Because α-ENaC knockout mice died of respiratory failure within 40 hours of birth and had increased lung wet weights, it was assumed that deletion of the α-ENaC gene disrupted airway liquid clearance. 11 However, these newborn mice also had weak inspiratory activity, 18 which likely contributed to respiratory failure, because respiratory function is a major determinant of lung aeration at birth. 24,25 Indeed, deletion of β- or γ-ENaC subunits doesn’t cause respiratory failure despite reducing ENaC activity sixfold. 18 Similarly, infants with gene mutations that markedly reduce ENaC activity (pseudoaldosteronism) do not have respiratory failure at birth. 26
At birth, liquid must partly fill the airways because air is unlikely to enter the distal airways before the head is delivered. As such, a significant amount of liquid must be cleared after birth, particularly from the distal airways. Recent studies indicate that postnatal respiratory activity plays the final and most significant role in airway liquid clearance 24,25 at birth.
Role of respiratory activity: Using phase contrast x-ray imaging, the entry of air into the lungs after birth has been imaged to study the temporal and spatial pattern of lung aeration. These studies demonstrate that lung aeration mostly (∼95%) occurs during inspiration in spontaneously breathing term newborn rabbits, with no liquid clearance occurring between breaths; videos can be viewed at http://www.neoresus.org.au/pages/LM1-7-Breathing.php#B _FirstBreaths and at http://www.fasebj.org/cgi/content/full/fj.07-8208com/DC1.25,26 Instead, the air–liquid interface tended to move proximally between breaths, reflecting liquid reentry into the airways and a gradual decline in functional residual capacity (FRC). As amiloride increases the rate of FRC decrease. 23 Na + reabsorption may assist in maintaining FRC by minimising liquid reentry into the airways between breaths. Lung aeration and FRC accumulation only occurred during inspirations, resulting in a stepwise increase in FRC with each breath. 24,25 The FRC increase per breath varied but was as high as 3 mL/kg in some newborns, resulting in an FRC of 1 5mL/kg after 5 breaths, over about 30 seconds. 24,25 Thus with an inspiratory time of about 0.3 seconds, these newborn rabbits cleared airway liquid at a rate of about 9 mL/kg/sec (or ∼32 L/kg/hr) during inspiration, which is considerably greater than what can be achieved with adrenaline (maximum of 10 mL/kg/hr). 14-16
Inspiration-induced airway liquid clearance is thought to result from an increase in the transepithelial pressure gradient. 24,25 This gradient is generated by inspiratory muscles, which reduce intrapleural and interstitial tissue pressures by expanding the chest wall. This produces a pressure gradient across the airway wall, between the interstitial tissue and airway lumen, as well as between the upper and lower airways. As a result, liquid is driven from the proximal into the distal airways from where it is cleared across the distal airway wall into the surrounding interstitial tissue space.
How do glucocorticoids mature the developing lung, and how do these changes allow efficient gas exchange at birth?
Circulating fetal cortisol concentrations increase markedly before birth, which is thought to structurally mature the lungs in preparation for air breathing. These changes include thinning of the lung mesenchymal tissue, increasing tissue compliance, promoting development of the vascular bed and increasing the differentiation of alveolar epithelial cells. Experimentally manipulating the levels of fetal cortisol alters lung structural development. Whereas fetal adrenalectomy or hypophysectomy delays lung development, infusion of cortisol to the fetus accelerates the changes in lung development. Mice that have had the glucocorticoid receptor deleted die at birth because of respiratory failure and have lungs that are structurally immature. This is caused by a continued proliferation of the lung mesenchymal cells and altered extracellular matrix remodelling, preventing the normal thinning of the lung tissue and the air–blood barrier. Thus gas exchange cannot be efficiently performed. Because of their role in promoting lung maturation, synthetic glucocorticoids are given clinically to women at risk for preterm labour, which has led to a marked increase in the survival of premature babies.
te Pas A.B., Davis P.G., Hooper S.B., Morley C.J. From liquid to air: breathing after birth. J Pediatr of cs . 2008;152:607–611.
Hooper S.B., te Pas A.B., Lewis R.A., Morley C.J. Establishing functional residual capacity at birth. NeoReviews . 2010;11:474–483.
Hooper S.B., Siew M.L., Kitchen M.J., te Pas A.B. Establishing functional residual capacity in the non-breathing infant. Semin Fetal Neonatal Med . 2013;18(6):336–343.
Olver R.E., Walters D.V., Wilson M. Developmental regulation of lung liquid transport. Annu Rev Physiol . 2004;66:77–101.
Berger P.J., Kyriakides M.A., Smolich J.J., et al. Massive decline in lung liquid before vaginal delivery at term in the fetal lamb. Am J Obstet Gynecol. 1998;178:223–227.
Lines A., Hooper S.B., Harding R. Lung liquid production rates and volumes do not decrease before labor in healthy fetal sheep. J Appl Physiol . 1997;82:927–932.
Harding R., Hooper S.B., Dickson K.A. A mechanism leading to reduced lung expansion and lung hypoplasia in fetal sheep during oligohydramnios. Am J Obstet Gynecol . 1990;163:1904–1913.
Jain L., Dudell G.G. Respiratory transition in infants delivered by cesarean section. Semin Perinatol . 2006;30:296–304.
Vyas H., Field D., Milner A.D., Hopkin I.E. Determinants of the first inspiratory volume and functional residual capacity at birth. Pediatr Pulmonol . 1986;2:189–193.
Bland R.D. Loss of liquid from the lung lumen in labor: more than a simple ‘squeeze’. Am J Physiol Lung Cell Mol Physiol . 2001;280:L602–L605.
Hummler E., Planès C. Importance of ENaC-mediated sodium transport in alveolar fluid clearance using genetically-engineered mice. Cell Physiol Biochem . 2010;25(1):63–70.
O’Donnell C.P., Kamlin C.O., Davis P.G., Morley C.J. Crying and breathing by extremely preterm infants immediately after birth. J Pediatr . 2010;156:846–847.
te Pas A.B., Davis P.G., Kamlin C.O., et al. Spontaneous breathing patterns of very preterm infants treated with continuous positive airway pressure at birth. Pediatr Res . 2008;64:281–285.
Olver R.E., Ramsden C.A., Strang L.B., Walters D.V. The role of amiloride-blockable sodium transport in adrenaline-induced lung liquid reabsorption in the fetal lamb. J Physiol. 1986;376:321–340.
Hooper S.B., Harding R. Effects of beta-adrenergic blockade on lung liquid secretion during fetal asphyxia. Am J Physiol . 1989;257:R705–R710.
Walters D.V., Olver R.E. The role of catecholamines in lung liquid absorption at birth. Pediatr Res . 1978;12:239–242.
Wallace M.J., Hooper S.B., Harding R. Regulation of lung liquid secretion by arginine vasopressin in fetal sheep. Am J Physiol . 1990;258:R104–R111.
Lagercrantz H., Bistoletti P. Catecholamine release in newborn-infant at birth. Pediatr Res. 1973;11:889–893.
Mortola J.P. Dynamics of breathing in newborn mammals. Physiol Rev . 1987;67(1):187–243.
Karlberg P., Cherry R.B., Escardo F.E., Koch G. Respiratory studies in newborn infants. II. Pulmonary ventilation and mechanics of breathing in first minutes of life, including onset of respiration. Acta Paediatr. 1962;51:121–136.
Dawson J.A., Kamlin C.O., Wong C., et al. Changes in heart rate in the first minutes after birth. Arch Dis Child Fetal Neonatal Ed . 2010;95:F177–F181.
Jain L., Eaton D.C. Physiology of fetal lung fluid clearance and the effect of labor. Semin Perinatol. 2006;30:34–43.
Siew M.L., Wallace M.J., Allison B.J., et al. The role of lung inflation and sodium transport in airway liquid clearance during lung aeration in newborn rabbits. Pediatr Res . 2013;73:443–449.
Hooper SB, Kitchen MJ, Wallace MJ, et al. Imaging lung aeration and lung liquid clearance at birth. FASEB J . 2007;21:3329–3337. 3.
Siew M.L., Wallace M.J., Kitchen M.J., et al. Inspiration regulates the rate and temporal pattern of lung liquid clearance and lung aeration at birth. J Appl Physiol. 2009;106:1888–1895.
Bonny O, Rossier BC. Disturbances of Na/K balance: pseudohypoaldosteronismrevisited. J Am Soc Nephrol . 2002;13:2399–414.
How many kidneys have most people had over their life course?
Six, consisting of the two pronephroi and two mesonephroi that mostly disappear during fetal development and two metanephroi that develop into the adult kidneys.
What is the single most important factor that determines kidney function?
Nephron number ultimately determines renal function, although prerenal, renal and postrenal issues can all affect apparent function.
Name three established causes of congenital anomalies of the kidney and urinary tract (CAKUT).
Genetic defects, obstruction and teratogens and diet have all been linked to CAKUT pathogenesis, although most cases do not have a clear cause, suggesting multifactorial effects.
What condition should be considered in a fetus with CAKUT and a family history of diabetes?
Mutations in the TCF2 gene, encoding the transcription factor hepatocyte nuclear factor-1β, are seen in the renal cysts and diabetes syndrome. Confusingly, this is not always associated with cysts because it accounts for up to one third of antenatal presentations with ‘bright’ kidneys and 10% of CAKUT cases overall in a spectrum also, including renal hypoplasia and agenesis.
Describe the secondary changes that occur to a fetus during a period of intrauterine retention.
After an intrauterine death, there may be a period of intrauterine retention of the fetus before delivery. During this period, a series of changes occur, known as maceration. Signs of maceration include blistering and slippage of the skin, generalised discolouration and increased laxity of the skull bones. The fetal birth weight also decreases with increasing intrauterine interval ( eFig. 13.1 ). 1
Internal changes during retention include changes in the weight of organs, with particularly prominent loss of mass from visceral organs such as the liver, 2 which affect the brain-to-liver weight ratio. The fetal brain-to-liver weight ratio may provide useful information regarding intrauterine growth status at time of death; however, adjustments should be made for weight changes to avoid mistaking postmortem fetal weight loss caused by maceration for intrauterine growth restriction.
 and adjusted birth weight in 308 stillbirths at 24 gestational weeks or later for whom well‐documented IUI data were provided. There was a significant linear relationship between increasing IUI and reducing delta birth weight (y = −0.0753x – 0.581); P <.0001) ( A ). This relationship persisted even when only data with IUI less than 12 days were evaluated; ( n = 296; y = −0.2081x – 0.3174) ( B ). (Reproduced with permission from Man J, Hutchinson JC, Ashworth M, et al. Effects of intrauterine retention and postmortem interval on body weight following intrauterine death: implications for assessment of fetal growth restriction at autopsy. Ultrasound Obstet Gynecol 48(5):574–578, 2016.)")
What postmortem imaging techniques are available as part of less invasive autopsy?
The range of imaging modalities and techniques include postmortem skeletal survey (i.e. radiography), postmortem ultrasonography (PMUS), postmortem magnetic resonance imaging (PMMRI), postmortem computed tomography (PMCT), and micro-focus computed tomography (micro CT) ( eFigs. 13.2 to 13.4 ).
PMUS is an operator-dependent technique that can provide good soft tissue differentiation with real-time imaging.
PMMR sequences also allows for good soft tissue differentiation and can provide information on bony cartilage and bone marrow changes. When three-dimensional (3D) volumetric imaging sequences have been obtained, the images can be reconstructed in nonanatomical planes. The limitations to this technique include loss of resolution for smaller fetuses and poor ossified bony detail, which can be of particular interest when a known musculoskeletal disorder is present.
PMCT using conventional imaging methods (i.e. medical CT imaging as for live patients) provides little to no soft tissue differentiation, with poor visualisation of internal organs but better assessment of the ossified bone compared with ultrasound or MRI techniques. Volume rendered imaging techniques enable a 3D skeleton of the deceased to be reconstructed and can allow for improved assessment of some fractures (particularly useful when differentiating skull fractures from sutures). Although intravascular contrast enhanced PMCT methods exist, this is not currently widely use in the perinatal paediatric setting.
Postmortem micro CT is the newest technique of all modalities mentioned here and is currently available in only a few centres worldwide. It can achieve high-resolution imaging down to a similar scale as light microscopy and provide information on the bony micro-architecture (when used without contrast) and excellent soft tissue differentiation (when used with contrast). Limitations include limited availability and the ability to image whole fetuses conforming below a certain size (because of the image detector dimensions).
 ( A ) and after a volume rendered reconstruction of the ossified skeleton from conventional computed tomography imaging of the whole body ( B ).")
, T2-weighted MRI sequences ( B ) and noncontrast computed tomography (CT) ( C ). Note the difference in brain parenchymal and ventricular anatomy on both on ultrasonography and MRI compared with the homogenous density of the intracranial structures seen on the CT imaging, where it is impossible to differentiate any intracranial structures apart from the skull encasing them.")
, T2-weighted magnetic resonance imaging (MRI) sequences ( B ) and noncontrast computed tomography (CT) ( C ). The ultrasound and MRI images allow for visualisation of the kidneys (white arrows) ; however, artefact from the vertebral bodies obscure views of the spinal canal, which is better seen on the MRI study (red arrows) . On CT imaging, the internal organs are not appreciated, although the appearances of the vertebral ossification centres are best seen with this modality.")
Describe the role of placental examination in the investigation of an intrauterine fetal death.
After an intrauterine death or a complicated delivery, the placenta should always be sent for histopathological examination. As the placenta is considered to be the mother’s tissue, consent for an autopsy is not required for placental examination alone, although placental examination also forms an integral part of the perinatal autopsy process. Features identified within the placenta (either at gross examination or at microscopic examination of placental samples taken by the pathologist) may help to explain the cause of a baby born in unexpectedly poor condition and may be medicolegally relevant to the defense of medical care provided to the mother during the peripartum period.
One recent large study of 1064 intrauterine fetal deaths found that around one third of cases in which the placenta was submitted for examination had the cause of death assigned to abnormalities of the placenta, cord or membranes. 3 Additionally, a minority of cases are associated with placental pathologies that may recur in subsequent pregnancies (e.g. fetal thrombotic vasculopathy, chronic histiocytic intervillositis and high-grade villitis of unknown aetiology). The significance of placental lesions is still the subject of much debate, and when the cause of death is unknown, placental lesions of unknown significance may be present.
Placental pathologies therefore represent the single largest cause of intrauterine fetal death, and placental histologic examination provides the highest yield of all aspects of investigations after death, including the physical autopsy and histopathology of the body viscera. 3
Man J., Hutchinson J.C., Ashworth M., et al. Effects of intrauterine retention and postmortem interval on body weight following intrauterine death: implications for assessment of fetal growth restriction at autopsy. Ultrasound Obstet Gynecol . 2016;48(5):574–578.
Man J., Hutchinson J.C., Ashworth M., et al. Organ weights and ratios for postmortem identification of fetal growth restriction: utility and confounding factors. Ultrasound Obstet Gynecol. 2016;48(5):585–590.
Man J., Hutchinson J.C., Heazell A.E., et al. Stillbirth and intrauterine fetal death: role of routine histopathological placental findings to determine cause of death. Ultrasound Obstet Gynecol . 2016;48(5):579–584.
What is ‘randomisation’ as in a randomised clinical trial?
Randomisation is the process by which consenting participants in a trial are randomly allocated into either the treatment (drug) or placebo arm of a two arm trial to assess the treatment efficacy. A well-implemented randomisation ensures that the participants in the two groups are well balanced (at baseline) as regards their characteristics, the study will be least biased, and when done well, will produce the highest level of causal evidence of treatment efficacy.
When is a randomised clinical trial applicable?
Randomised trials are applicable when the intent is to assess treatment efficacy of a new drug with an existing treatment regimen or a placebo, when there is a perceived benefit to testing a new drug or a therapy, when it is ethical to subject participants to either the drug (for safety concerns) or a placebo (for concerns of withholding treatment to really sick patients) and when there is availability of resources to carry out the trial (time and money).
What is the underlying philosophy of a meta-analysis?
Meta-analysis is a statistical methodology to combine results (or data) from several seemingly similar (or homogenous) studies to derive a single measure of association. An implicit goal, one that often gets overlooked in a meta-analysis, is its ability to uncover interstudy heterogeneity. This measure of heterogeneity will be helpful to design other future studies by avoiding the potential sources of heterogeneity that gets uncovered in a meta-analysis.
From what point in gestation does the future child have interests that should be taken into account in decision making about fetal treatment?
The interests of the future child count from conception.
Does the administration of pain relief during fetal surgery only serve the interests of the fetus as fetus or also those of the future child?
There is evidence that experiences of stress or pain will be ‘remembered’ by the nervous system in later life, so taking these measures also serves the interest of the future child.
If fetal treatment for a lethal condition saves the life of the fetus at the unavoidable price of bringing a child into the world with serious disabilities, to what extent is that morally problematic?
As long as the child has a life worth living, he or she will not have been harmed. However, the outcome may still be regarded as morally problematic because of other reasons (e.g. for bringing a harmful state into the world, for emotionally or financially burdening the child’s family).
If fetal neurocognitive treatment for Down syndrome can be shown to lead to a marked improvement in cognitive functioning for people with that condition, there may still be a concern that this would not really benefit them. What might this concern be?
It is possible that a partial improvement in cognitive functioning will make persons with Down syndrome only more aware of not being able to fully participate in society or to realise the professional and reproductive options open to others.
What are the basic statistical parameters used to evaluate a screening test?
Become a Clinical Tree membership for Full access and enjoy Unlimited articles
If you are a member. Log in here