Physical Address
304 North Cardinal St.
Dorchester Center, MA 02124

The regulation of hydrogen ion (H + ) balance is similar in some ways to the regulation of other ions in the body. For example, there must be a balance between the intake or production of H + and the net removal of H + from the body to achieve homeostasis. And, as is true for other ions, the kidneys play a key role in regulating H + removal from the body. However, precise control of extracellular fluid H + concentration involves much more than the simple elimination of H + by the kidneys. Multiple acid–base buffering mechanisms involving the blood, cells, and lungs also are essential in maintaining normal H + concentrations in extracellular and intracellular fluids.
In this chapter, we consider the mechanisms that regulate H + concentration, with special emphasis on renal H + secretion and renal reabsorption, production, and excretion of bicarbonate ions (HCO 3 − ), one of the key components of acid–base control systems in the body fluids.
Precise H + regulation is essential because the activities of almost all enzyme systems in the body are influenced by H + concentration. Therefore, changes in H + concentration alter virtually all cell and body functions.
Compared with other ions, the H + concentration of the body fluids normally is kept at a low level. For example, the concentration of sodium in extracellular fluid (142 mEq/L) is about 3.5 million times as great as the normal concentration of H + , which averages only 0.00004 mEq/L. Equally important, the normal variation in H + concentration in extracellular fluid is only about one millionth as great as the normal variation in sodium ion (Na + ) concentration. Thus, the precision with which H + is regulated emphasizes its importance to the various cell functions.
A hydrogen ion is a single free proton released from a hydrogen atom. Molecules containing hydrogen atoms that can release hydrogen ions in a solution are referred to as acids . An example is hydrochloric acid (HCl), which ionizes in water to form hydrogen ions (H + ) and chloride ions (Cl − ). Likewise, carbonic acid (H 2 CO 3 ) ionizes in water to form H + and bicarbonate ions (HCO 3 − ).
A base is an ion or a molecule that can accept an H + . For example, HCO 3 − is a base because it can combine with H + to form H 2 CO 3 . Likewise, HPO 4 = is a base because it can accept an H + to form H 2 PO 4 − . The proteins in the body also function as bases because some of the amino acids that make up proteins have net negative charges that readily accept H + . The protein hemoglobin in the red blood cells and proteins in the other cells of the body are among the most important of the body’s bases.
The terms base and alkali are often used synonymously. An alkali is a molecule formed by the combination of one or more of the alkaline metals—such as sodium, potassium, and lithium—with a highly basic ion such as a hydroxyl ion (OH − ). The base portion of these molecules reacts quickly with H + to remove it from solution and are, therefore, typical bases. For similar reasons, the term alkalosis refers to the excess removal of H + from the body fluids, in contrast to the excess addition of H + , which is referred to as acidosis.
A strong acid, such as HCl, rapidly dissociates and releases especially large amounts of H + in solution. Weak acids such as H 2 CO 3 are less likely to dissociate their ions and, therefore, release H + with less vigor. A strong base is one that reacts rapidly and strongly with H + and, therefore, quickly removes H + from a solution. A typical example is OH − , which reacts with H + to form water (H 2 O). A typical weak base is HCO 3 − because it binds with H + much more weakly than OH − . Most acids and bases in the extracellular fluid that are involved in normal acid–base regulation are weak acids and bases. The most important ones that we discuss are carbonic acid (H 2 CO 3 ) and HCO 3 − base.
The blood H + concentration is normally maintained within tight limits around a normal value of about 0.00004 mEq/L (40 nEq/L). Normal variations are only about 3 to 5 nEq/L but, under extreme conditions, the H + concentration can vary from as low as 10 nEq/L to as high as 160 nEq/L without resulting in death.
Because H + concentration normally is low, and because these small numbers are cumbersome, it is customary to express H + concentration on a logarithm scale using pH units. pH is related to the actual H + concentration by the following formula (H + concentration [H + ] is expressed in equivalents per liter):
For example, normal [H + ] is 40 nEq/L (0.00000004 Eq/L). Therefore, the normal pH is as follows:
From this formula, one can see that pH is inversely related to the H + concentration; therefore, a low pH corresponds to a high H + concentration, and a high pH corresponds to a low H + concentration.
The normal pH of arterial blood is 7.4, whereas the pH of venous blood and interstitial fluids is about 7.35 because of the extra amounts of carbon dioxide (CO 2 ) released from the tissues to form H 2 CO 3 in these fluids ( Table 31-1 ). Because the normal pH of arterial blood is 7.4, a person is considered to have acidemia when the pH falls significantly below this value and alkalemia when the pH rises above 7.4. The lower limit of pH at which a person can live more than a few hours is about 6.8, and the upper limit is about 8.0.
| H + Concentration (mEq/L) | pH | |
|---|---|---|
| Extracellular Fluid | ||
|
|
|
|
|
|
|
|
|
| Intracellular fluid | 1 × 10 −3 to 4 × 10 −5 | 6.0–7.4 |
| Urine | 3 × 10 −2 to 1 × 10 −5 | 4.5–8.0 |
| Gastric HCl | 160 | 0.8 |
Intracellular pH usually is slightly lower than plasma pH because cell metabolism produces acid, especially H 2 CO 3 . Depending on the type of cells, the pH of intracellular fluid has been estimated to range between 6.0 and 7.4. Hypoxia of the tissues and poor blood flow to the tissues can cause acid accumulation and decreased intracellular pH. The terms acidosis and alkalosis describe the processes that lead to acidemia and alkalemia, respectively.
The pH of urine can range from 4.5 to 8.0, depending on the acid–base status of the extracellular fluid. As discussed later, the kidneys play a major role in correcting abnormalities of extracellular fluid H + concentration by excreting acids or bases at variable rates.
An extreme example of an acidic body fluid is the HCl secreted into the stomach by the oxyntic (parietal) cells of the stomach mucosa, as discussed in Chapter 65 . The H + concentration in these cells is about 4 million times greater than the hydrogen concentration in blood, with a pH of 0.8. In the remainder of this chapter, we discuss the regulation of extracellular fluid H + concentration.
Three primary systems regulate the H + concentration in the body fluids: (1) the chemical acid–base buffer systems of the body fluids , which immediately combine with an acid or a base to prevent excessive changes in H + concentration; (2) the respiratory center , which regulates the removal of CO 2 (and, therefore, H 2 CO 3 ) from the extracellular fluid; and (3) the kidneys , which can excrete acidic or alkaline urine, thereby readjusting the extracellular fluid H + concentration toward normal during acidosis or alkalosis.
When there is a change in H + concentration, the buffer systems of the body fluids react within seconds to minimize these changes. Buffer systems do not eliminate H + from or add H + to the body but only keep them tied up until balance can be re-established.
The second line of defense, the respiratory system , acts within a few minutes to eliminate CO 2 and, therefore, H 2 CO 3 from the body.
These first two lines of defense keep the H + concentration from changing too much until the more slowly responding third line of defense, the kidneys , can eliminate the excess acid or base from the body. Although the kidneys are relatively slow to respond compared with the other defenses, over a period of hours to several days, they are by far the most powerful of the acid–base regulatory systems.
A buffer is any substance that can reversibly bind H + . The general form of the buffering reaction is as follows:
In this example, a free H + combines with the buffer to form a weak acid (H buffer) that can either remain as an unassociated molecule or dissociate back to the buffer and H + . When the H + concentration increases, the reaction is forced to the right, and more H + binds to the buffer, as long as buffer is available. Conversely, when the H + concentration decreases, the reaction shifts toward the left, and H + is released from the buffer. In this way, changes in H + concentration are minimized.
The importance of the body fluid buffers can be quickly realized if one considers the low concentration of H + in the body fluids and the relatively large amounts of acids produced by the body each day. About 80 milliequivalents of H + is ingested or produced each day by metabolism, whereas the H + concentration of the body fluids normally is only about 0.00004 mEq/L. Without buffering, the daily production and ingestion of acids would cause lethal changes in the body fluid H + concentration.
The action of acid–base buffers can perhaps best be explained by considering the buffer system that is quantitatively the most important in the extracellular fluid—the bicarbonate buffer system.
The bicarbonate buffer system consists of a water solution that contains two ingredients: (1) a weak acid, H 2 CO 3 ; and (2) a bicarbonate salt, such as sodium bicarbonate (NaHCO 3 ).
H 2 CO 3 is formed in the body by the reaction of CO 2 with H 2 O:
This reaction is slow, and exceedingly small amounts of H 2 CO 3 are formed unless the enzyme carbonic anhydrase is present. This enzyme is especially abundant in the walls of the lung alveoli, where CO 2 is released; carbonic anhydrase is also present in the epithelial cells of the renal tubules, where CO 2 reacts with H 2 O to form H 2 CO 3 .
H 2 CO 3 ionizes weakly to form small amounts of H + and HCO 3 − :
The second component of the system, bicarbonate salt, occurs predominantly as NaHCO 3 in the extracellular fluid. NaHCO 3 ionizes almost completely to form HCO 3 − and Na + , as follows:
Now, putting the entire system together, we have the following:
Because of the weak dissociation of H 2 CO 3 , the H + concentration is extremely low.
When a strong acid such as HCl is added to the bicarbonate buffer solution, the increased H + released from the acid (HCl → H + + Cl − ) is buffered by HCO 3 − :
As a result, more H 2 CO 3 is formed, causing increased CO 2 and H 2 O production. From these reactions, one can see that H + from the strong acid HCl reacts with HCO 3 − to form the very weak acid H 2 CO 3 , which in turn forms CO 2 and H 2 O. The excess CO 2 greatly stimulates respiration, which eliminates the CO 2 from the extracellular fluid.
The opposite reactions take place when a strong base, such as sodium hydroxide (NaOH), is added to the bicarbonate buffer solution.
In this case, the OH − from the NaOH combines with H 2 CO 3 to form additional HCO 3 − . Thus, the weak base NaHCO 3 replaces the strong base NaOH. At the same time, the concentration of H 2 CO 3 decreases (because it reacts with NaOH), causing more CO 2 to combine with H 2 O to replace the H 2 CO 3 :
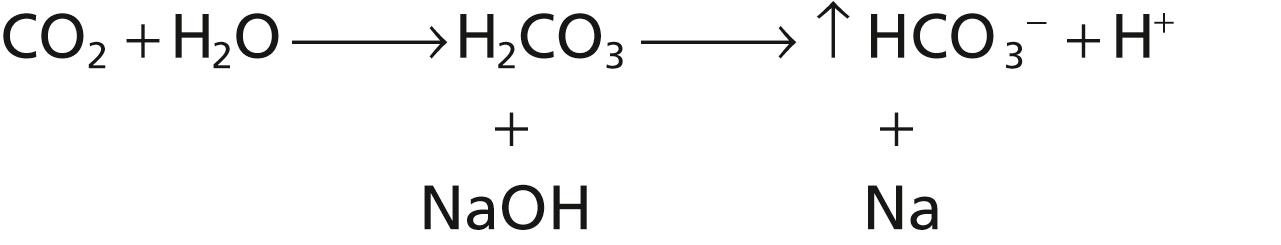
The net result, therefore, is a tendency for the CO 2 levels in the blood to decrease, but the decreased CO 2 in the blood inhibits respiration and decreases the rate of CO 2 expiration. The rise in blood HCO 3 − concentration that occurs is compensated for by increased renal excretion of HCO 3 − .
All acids, including H 2 CO 3 , are ionized to some extent. From mass balance considerations, the concentrations of H + and HCO 3 − are proportional to the concentration of H 2 CO 3 :
For any acid, the concentration of the acid relative to its dissociated ions is defined by the dissociation constant, K ′.
This equation indicates that in an H 2 CO 3 solution, the amount of free H + is equal to
The concentration of undissociated H 2 CO 3 cannot be measured in solution because it rapidly dissociates into CO 2 and H 2 O or into H + and HCO 3 − . However, the CO 2 dissolved in the blood is directly proportional to the amount of undissociated H 2 CO 3 . Therefore, Equation 2 can be rewritten as follows:
The dissociation constant (K) for Equation 3 is only about 1/400 of the dissociation constant (K′) of Equation 2 because the proportionality ratio between H 2 CO 3 and CO 2 is 1:400.
Equation 3 is written in terms of the total amount of CO 2 dissolved in solution. However, most clinical laboratories measure the blood CO 2 tension (P co 2 ) rather than the actual amount of CO 2 . Fortunately, the amount of CO 2 in the blood is a linear function of P co 2 multiplied by the solubility coefficient for CO 2 ; under physiological conditions, the solubility coefficient for CO 2 is 0.03 mmol/mm Hg at body temperature. This means that 0.03 millimole of H 2 CO 3 is present in the blood for each mm Hg P co 2 measured. Therefore, Equation 3 can be rewritten as
As discussed earlier, it is customary to express the H + concentration in pH units rather than in actual concentrations. Recall that pH = −log H + .
The dissociation constant (pK) can be expressed in a similar manner.
Therefore, we can express the H + concentration in Equation 4 in pH units by taking the negative logarithm of that equation, which yields the following:
Therefore,
Rather than work with a negative logarithm, we can change the sign of the logarithm and invert the numerator and denominator in the last term, using the law of logarithms, to yield the following:
For the bicarbonate buffer system, the pK is 6.1, and Equation 7 can be written as follows:
Equation 8 is the Henderson-Hasselbalch equation and, with it, one can calculate the pH of a solution if the molar concentration of HCO 3 − and the P co 2 are known.
From the Henderson-Hasselbalch equation, it is apparent that an increase in HCO 3 − concentration causes the pH to rise, shifting the acid–base balance toward alkalosis. An increase in P co 2 causes the pH to decrease, shifting the acid–base balance toward acidosis.
The Henderson-Hasselbalch equation, in addition to defining the determinants of normal pH regulation and acid–base balance in the extracellular fluid, provides insight into the physiological control of the acid and base composition of the extracellular fluid. As discussed later, the HCO 3 − concentration is regulated mainly by the kidneys, whereas the P co 2 in extracellular fluid is controlled by the rate of respiration. By increasing the rate of respiration, the lungs remove CO 2 from the plasma and, by decreasing respiration, the lungs elevate P co 2 . Normal physiological acid–base homeostasis results from the coordinated efforts of both the lungs and kidneys, and acid–base disorders occur when one or both of these control mechanisms are impaired, thus altering either the HCO 3 − concentration or P co 2 of extracellular fluid.
When disturbances of acid–base balance result from a primary change in extracellular fluid HCO 3 − concentration, they are referred to as metabolic acid–base disorders. Therefore, acidosis caused by a primary decrease in HCO 3 − concentration is termed metabolic acidosis , whereas alkalosis caused by a primary increase in HCO 3 − concentration is called metabolic alkalosis . Acidosis caused by an increase in P co 2 is called respiratory acidosis , whereas alkalosis caused by a decrease in P co 2 is termed respiratory alkalosis .
Figure 31-1 shows the changes in pH of the extracellular fluid when the ratio of HCO 3 − to CO 2 in extracellular fluid is altered. When the concentrations of these two components are equal, the right-hand portion of Equation 8 becomes the log of 1, which is equal to 0. Therefore, when the two components of the buffer system are equal, the pH of the solution is the same as the pK (6.1) of the bicarbonate buffer system. When base is added to the system, part of the dissolved CO 2 is converted into HCO 3 − , causing an increase in the ratio of HCO 3 − to CO 2 and increasing the pH, as is evident from the Henderson-Hasselbalch equation. When acid is added, it is buffered by HCO 3 − , which is then converted into dissolved CO 2 , decreasing the ratio of HCO 3 − to CO 2 and decreasing the pH of the extracellular fluid.
 are altered.")
From the titration curve in Figure 31-1 , several points are apparent. First, the pH of the system is the same as the pK when each of the components (HCO 3 − and CO 2 ) constitutes 50% of the total concentration of the buffer system. Second, the buffer system is most effective in the central part of the curve, where the pH is near the pK of the system. This phenomenon means that the change in pH for any given amount of acid or base added to the system is least when the pH is near the pK of the system. The buffer system is still reasonably effective for 1.0 pH unit on either side of the pK, which, for the bicarbonate buffer system, extends from a pH of about 5.1 to 7.1 units. Beyond these limits, the buffering power rapidly diminishes. And, when all the CO 2 has been converted into HCO 3 − , or when all the HCO 3 − has been converted into CO 2 , the system has no more buffering power.
The absolute concentration of the buffers is also an important factor in determining the buffer power of a system. With low concentrations of the buffers, only a small amount of acid or base added to the solution changes the pH considerably.
From the titration curve shown in Figure 31-1 , one would not expect the bicarbonate buffer system to be powerful, for two reasons. First, the pH of the extracellular fluid is about 7.4, whereas the pK of the bicarbonate buffer system is 6.1, which means that there is about 20 times as much of the bicarbonate buffer system in the form of HCO 3 − as in the form of dissolved CO 2 . For this reason, this system operates on the portion of the buffering curve where the slope is low, and the buffering power is poor. Second, the concentrations of the two elements of the bicarbonate system, CO 2 and HCO 3 − , are not high.
Despite these characteristics, the bicarbonate buffer system is the most powerful extracellular buffer in the body. This apparent paradox is due mainly to the fact that the two elements of the buffer system, HCO 3 − and CO 2 , are regulated, respectively, by the kidneys and lungs, as discussed later. As a result of this regulation, the pH of the extracellular fluid can be precisely controlled by the relative rate of removal and addition of HCO 3 − by the kidneys and rate of removal of CO 2 by the lungs.
Although the phosphate buffer system is not a major extracellular fluid buffer, it plays an important role in buffering renal tubular fluid and intracellular fluids.
The main elements of the phosphate buffer system are H 2 PO 4 − and HPO 4 = . When a strong acid such as HCl is added to a mixture of these two substances, the hydrogen is accepted by the base HPO 4 = and converted to H 2 PO 4 − :
The result of this reaction is that the strong acid, HCl, is replaced by an additional amount of a weak acid, NaH 2 PO 4 , and the decrease in pH is minimized.
When a strong base, such as NaOH, is added to the buffer system, the OH − is buffered by the H 2 PO 4 − to form additional amounts of HPO 4 = + H 2 O:
In this case, a strong base, NaOH, is traded for a weak base, Na 2 HPO 4 , causing only a slight increase in pH.
The phosphate buffer system has a pK of 6.8, which is not far from the normal pH of 7.4 in the body fluids, allows the system to operate near its maximum buffering power. However, its concentration in the extracellular fluid is low, at only about 8% of the concentration of the bicarbonate buffer. Therefore, the total buffering power of the phosphate system in the extracellular fluid is much less than that of the bicarbonate buffering system.
In contrast to its minor role as an extracellular buffer, the phosphate buffer is especially important in the tubular fluids of the kidneys for two reasons: (1) phosphate usually becomes greatly concentrated in the tubules, thereby increasing the buffering power of the phosphate system; and (2) the tubular fluid usually has a considerably lower pH than the extracellular fluid, bringing the operating range of the buffer closer to the pK (6.8) of the system.
The phosphate buffer system is also important in buffering intracellular fluid because the concentration of phosphate in this fluid is many times higher than in the extracellular fluid. Also, the pH of intracellular fluid is lower than that of extracellular fluid and, therefore, is usually closer to the pK of the phosphate buffer system compared with the extracellular fluid.
Proteins are among the most plentiful buffers in the body because of their high concentrations, especially in the cells. The pH of the cells, although slightly lower than in the extracellular fluid, nevertheless changes approximately in proportion to extracellular fluid pH changes. There is a slight diffusion of H + and HCO 3 − through the cell membrane, although these ions require several hours to reach equilibrium with the extracellular fluid, except for the rapid equilibrium that occurs in the red blood cells. CO 2 , however, can rapidly diffuse through all the cell membranes. This diffusion of the elements of the bicarbonate buffer system causes the pH in intracellular fluid to change when there are changes in extracellular pH. For this reason, the buffer systems in the cells help prevent changes in the pH of the extracellular fluid but may take several hours to become maximally effective.
In the red blood cell, hemoglobin (Hb) is an important buffer, as follows:
Approximately 60% to 70% of the total chemical buffering of the body fluids is inside the cells, and most of this buffering results from the intracellular proteins. However, except for the red blood cells, the slow rate at which H + and HCO 3 − move through the cell membranes often delays the maximum ability of the intracellular proteins to buffer extracellular acid–base abnormalities for several hours.
In addition to the high concentration of proteins in the cells, another factor that contributes to their buffering power is the fact that the pKs of many of these protein systems are fairly close to intracellular pH.
We have been discussing buffer systems as though they operate individually in the body fluids. However, they all work together because H + is common to the reactions of all these systems. Therefore, whenever there is a change in H + concentration in the extracellular fluid, the balance of all the buffer systems changes at the same time. This phenomenon is called the isohydric principle and is illustrated by the following formula:
K 1 , K 2 , and K 3 are the dissociation constants of three respective acids—HA 1 , HA 2 , HA 3 —and A 1 , A 2 , A 3 are the concentrations of the free negative ions that constitute the bases of the three buffer systems.
The implication of this principle is that any condition that changes the balance of one of the buffer systems also changes the balance of all the others because the buffer systems actually buffer one another by shifting H + back and forth between them.
The second line of defense against acid–base disturbances is control of extracellular fluid CO 2 concentration by the lungs. An increase in ventilation eliminates CO 2 from extracellular fluid, which, by mass action, reduces the H + concentration. Conversely, decreased ventilation increases CO 2 and H + concentrations in the extracellular fluid.
CO 2 is formed continually in the body by intracellular metabolic processes. After it is formed, it diffuses from the cells into the interstitial fluids and blood, and the flowing blood transports it to the lungs, where it diffuses into the alveoli and then is transferred to the atmosphere by pulmonary ventilation. About 1.2 mol/L of dissolved CO 2 is normally in the extracellular fluid, corresponding to a P co 2 of 40 mm Hg.
If the rate of metabolic formation of CO 2 increases, the P co 2 of the extracellular fluid is likewise increased. Conversely, a decreased metabolic rate lowers the P co 2 . If the rate of pulmonary ventilation is increased, CO 2 is blown off from the lungs, and the P co 2 in the extracellular fluid decreases. Therefore, changes in pulmonary ventilation or the rate of CO 2 formation by the tissues can change the extracellular fluid P co 2 .
If the metabolic formation of CO 2 remains constant, the only other factor that affects P co 2 in extracellular fluid is the rate of alveolar ventilation. The higher the alveolar ventilation, the lower the P co 2 . As discussed previously, when CO 2 concentration increases, the H 2 CO 3 concentration and H + concentration also increase, thereby lowering extracellular fluid pH.
Figure 31-2 shows the approximate changes in blood pH that are caused by increasing or decreasing the alveolar ventilation rate. Note that increasing alveolar ventilation to about twice normal raises extracellular fluid pH by about 0.23. If the pH of the body fluids is 7.40 with normal alveolar ventilation, doubling the ventilation rate raises the pH to about 7.63. Conversely, a decrease in alveolar ventilation to one-fourth normal reduces the pH by 0.45. That is, if the pH is 7.4 at a normal alveolar ventilation, reducing the ventilation to one-fourth normal reduces the pH to 6.95. Because the alveolar ventilation rate can change markedly, from as low as 0 to as high as 15 times normal, one can easily understand how much the pH of the body fluids can be changed by the respiratory system.

Not only does the alveolar ventilation rate influence H + concentration by changing P co 2 of the body fluids, but the H + concentration affects the rate of alveolar ventilation. Thus, Figure 31-3 shows that alveolar ventilation rate increases four to five times normal as pH decreases from the normal value of 7.4 to the strongly acidic value of 7.0. Conversely, a rise in plasma pH above 7.4 causes a decrease in the ventilation rate. The change in ventilation rate per unit pH change is much greater at reduced levels of pH (corresponding to elevated H + concentration) compared with increased levels of pH. The reason for this is that as the alveolar ventilation rate decreases, as a result of an increase in pH (decreased H + concentration), the amount of oxygen added to the blood decreases, and the partial pressure of oxygen (P o 2 ) in the blood also decreases, which stimulates the ventilation rate. Therefore, the respiratory compensation for an increase in pH is not nearly as effective as the response to a marked reduction in pH.

Become a Clinical Tree membership for Full access and enjoy Unlimited articles
If you are a member. Log in here